


En este artículo discutimos investigaciones recientes relacionadas a los mecanismos neurobiológicos subyacentes al Trastorno por Déficit Atencional e Hiperactividad, (TDAH) en particular la dinámica de señalización dopaminérgica y la llamada red por defecto, que consiste en patrones de actividad que se generan durante el reposo. Ambos tipos de fenómenos han sido asociados al TDAH y aquí proponemos una relación entre ambas dinámicas, y cómo ésta puede estar afectada en el TDAH.
In this article we discuss recent findings on the neurobiological mechanisms underlying Attentional Deficit Hyperactivity Disorder (ADHD); specifically the dynamics of dopaminergic signaling and the default mode network, consisting of activity patterns generated during the resting state. Both phenomena have been related to ADHD, and we propose here a relationship between both dynamics, and how this can be affected in ADHD.
El trastorno por déficit atencional e hiperactividad (TDAH) es el trastorno neuropsiquiátrico más común en la infancia. Contiene tres elementos clínicos cardinales: hiperactividad, impulsividad e inatención. Esta condición es altamente heredable, y se da con mayor frecuencia en hombres, en una proporción 4:1 (especialmente el tipo hiperactivo/ impulsivo). Sin embargo, hay discusión acerca de si el tipo puramente inatento es subestimado clínicamente, el cual podría ser más frecuente en niñas que en niños (1, 2). El TDAH está siendo crecientemente reconocido como un trastorno que afecta toda la vida; un alto porcentaje de los niños con TDAH mantiene esta condición en la edad adulta, aunque se ha observado que los síntomas de hiperactividad tienden a disminuir con la edad (3). Por el contrario, los síntomas de inatención tienden a mantenerse a lo largo de la vida, lo cual también se ajusta al hecho que en adultos la prevalencia de mujeres con TDAH parece ser más alta que en niños.
Una interpretación ampliamente aceptada es que subyacente al TDAH existe una alteración en los mecanismos de control cognitivo y conductual. En los últimos años se han generado grandes avances en la comprensión de los procesos neurobiológicos involucrados en dichos mecanismos, parte de los cuales se ha proyectado en el estudio de trastornos como el TDAH. En este breve artículo, se pondrán en contexto algunos de dichos avances, en particular respecto de la teoría dopaminérgica del TDAH, que indica que existe una disfunción de este neurotransmisor como base para su sintomatología. Además, se discutirán algunos estudios recientes de imaginología acerca de la dinámica cerebral durante el desempeño de tareas cognitivas, y sus alteraciones en el TDAH y otras condiciones neuropsiquiátricas. Fundamentalmente, apuntará a la existencia de redes neuronales que son en parte antagónicas y cuyas dinámicas y mutuos balances permiten la generación de conductas adecuadas en contextos específicos; y cómo éstas dinámicas pueden verse distorsionadas en el TDAH. El objetivo de este trabajo es actualizar parte de la información en investigación en TDAH, en particular desde una perspectiva del análisis de redes neuronales en el contexto de la generación de fenómenos cognitivos y últimamente la conducta.
La hipótesis dopaminérgica del TDAHEl TDAH es tratado primariamente con estimulantes como anfetaminas o metilfenidato, que activan el sistema catecolaminérgico (dopamina, epinefrina y norepinefrina), beneficiando a cerca de un 80% de los pacientes. En segunda opción se usa la atomoxetina, una droga no estimulante que actúa primariamente sobre el sistema noradrenérgico (4). El éxito de los estimulantes en el tratamiento del TDAH ha llevado a proponer la hipótesis dopaminérgica del TDAH, según la cual un déficit en la neurotransmisión dopaminérgica (y más en general, catecolaminérgica) en estos sujetos podría dar cuenta de gran parte de su sintomatología (5). La disfunción de este neurotransmisor afecta el funcionamiento de circuitos fronto-estriatales y fronto-cerebelares involucrados en la predicción de eventos y en la función inhibitoria, y también involucra el circuito fronto-amigdalino que le asigna un contenido emocional a dichos eventos (6). En esta línea, se ha distinguido entre los circuitos “fríos”, o puramente cognitivos, y los circuitos “calientes” involucrados en el control emocional, como dos elementos cuya alteración contribuye a la diversidad de la sintomatología observada en TDAH (6). Es necesario notar que evidencias recientes también apuntan a la participación de otros sistemas de neurotransmisores además de las catecolaminas, como la serotonina y la acetilcolina en la sintomatología de esta condición (7).
La hipótesis dopaminérgica ha sido parcialmente apoyada por la existencia de asociaciones relativamente débiles, pero consistentes, entre el TDAH y polimorfismos de genes asociados al sistema dopaminérgico. Nuestros propios estudios indican que en la población chilena, la coexistencia de dos alelos de riesgo para TDAH (uno correspondiente al receptor dopaminérgico DRD4, y el otro al transportador de dopamina DAT1), confiere un riesgo significativamente superior de desarrollar el TDAH que la suma de ambos alelos por separado (2, 8).
Si bien la hipótesis dopaminérgica ha sido ampliamente aceptada, aún no tiene un apoyo experimental directo (9) y existen opiniones fuertemente disidentes (10, 11). Algunos estudios han reportado una mayor densidad del transportador de dopamina en pacientes que aún no han sido tratados, la cual se normaliza después de la medicación con estimulante (12, 13). Dicho aumento de densidad ha sido interpretado como un estado hipodopaminérgico basal, lo cual resulta en la regulación positiva del transportador. En esta línea, hay estudios que sugieren una disminución de los receptores tipo D2 en pacientes sin tratamiento, lo cual se correlacionaría con los síntomas de inatención (14, 15). Sin embargo, no es claro si esto refleja una disminución real de la actividad dopaminérgica, o resulta de una disminución de receptores debida a la elevación del tono dopaminérgico basal (7). Otros reportes no han encontrado diferencias, o incluso una disminución de la densidad del transportador (16, 17). Gran parte de las discordancias en estos estudios puede relacionarse al radioligando específico que se usa en cada caso, ya que muchas veces dichos fármacos muestran reacción cruzada con otros transportadores.
Balance en la cuerda flojaNuestro grupo se ha dedicado a estudiar las bases neurobiológicas del TDAH desde hace cerca de 15 años, focalizándonos en los aspectos neurocognitivos y genéticos de esta condición. Nuestra hipótesis básica es que más que un déficit dopaminérgico generalizado, en el TDAH existe un desbalance entre distintos modos de neurotransmisión catecolaminérgica (dopamina y norepinefrina), pero también posiblemente de otros neurotransmisores (7, 18, 19). Una de las funciones primordiales de las catecolaminas tiene que ver con la regulación de la conducta orientada a metas, que consiste en la capacidad de responder a estímulos que predicen la aparición de un evento, y la subsecuente orientación de la conducta en relación a dicho evento. Existen dos formas de neurotransmisión dopaminérgica (y noradrenérgica) (20). La primera es la Liberación Fásica, que consiste en fuertes pero transientes andanadas de actividad dopaminérgica, ligadas a la aparición de estímulos relevantes; ésta es mediada principalmente por receptores excitatorios tipo D1 (en el caso de dopamina). La llamada Liberación Tónica consiste en la mantención de niveles basales de dopamina liberada en el espacio extracelular a lo largo del tiempo, y es mediada por receptores tipo D2. La actividad fásica se asocia a la aparición de estímulos “salientes”, o motivantes; en tanto que la actividad tónica tiene una función bastante más compleja, que tiene que ver con una dinámica basal, en reposo; pero también su aumento gradual se relaciona con el proceso de anticipación a un evento, con la mantención del alerta en el mediano plazo, con la actualización de la información según el contexto, y con fenómenos más generales de plasticidad neuronal. En otras palabras, la liberación fásica pone en marcha el sistema conductual, pero la mantención de éste en el mediano y largo plazo depende de aumentos graduales en la liberación tónica. Ahora, aunque un leve aumento en la actividad tónica permite mantener la representación del objetivo en mente y adaptarse a cambios contextuales, niveles inapropiados (muy altos o muy bajos) de actividad tónica en un contexto determinado pueden producir distractibilidad, y si son demasiado altos, ansiedad que se traduce en inquietud (18, 19). Según nosotros, en el TDAH existiría un desbalance entre estas dos formas de señalización, lo cual puede generar estados de alta impulsividad (debido a un énfasis en la liberación fásica; 20), o estados de alta distractibilidad y “desconexión” con el ambiente, o incluso hiperactividad no dirigida causada por la ansiedad (debido a alteraciones en la liberación tónica) (para más detalles, ver 7, 18, 19).
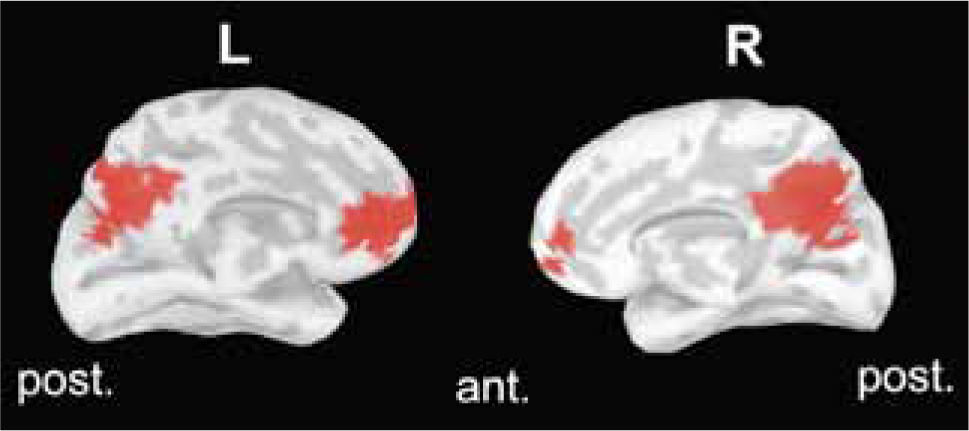
El cerebro en reposoEn 2001, usando resonancia magnética funcional (RMf) en humanos, Marcus Raichle y sus colaboradores publicaron un seminal artículo en el cual evidenciaron una verdadera red neuronal que involucraba regiones mediales en los aspectos anteriores y posteriores del hemisferio cerebral (la corteza cingulada posterior o precúneo, y la corteza frontal polar; ver Fig. 1). Lo más destacable de esta red era que su máxima actividad se producía en estados de reposo, donde las personas están divagando mentalmente con los ojos abiertos, sin tener ninguna tarea que realizar; pero su actividad disminuye fuertemente una vez que el sujeto se involucra en una tarea cognitiva determinada, cualquiera que ésta sea (21).
 y la región frontal medial (anterior). Estas regiones tienden a desactivarse cuando el sujeto se involucra en una tarea cognitiva, al contrario de lo que sucede en regiones de la red de enganche (no mostradas). L, izquierda (Left); R, derecha (Right).")
Esta figura muestra la red por defecto en una vista medial del cerebro humano, usando la metodología de la solución inversa en EEG. En rojo, el patrón de activación observado durante el reposo, donde se involucra el precúneo o cín-gulo posterior (región posterior) y la región frontal medial (anterior). Estas regiones tienden a desactivarse cuando el sujeto se involucra en una tarea cognitiva, al contrario de lo que sucede en regiones de la red de enganche (no mostradas). L, izquierda (Left); R, derecha (Right).
Estas áreas conforman la llamada red por defecto (DMN, o Default-Mode Network), y participan en fenómenos como la introspección, la actividad autoreferente y en general, en la detección de estados internos. Notoriamente, la región medial posterior o precúneo es particularmente sensible a la aparición de estímulos periféricos distractores. Por otro lado, las regiones asociadas al desempeño o “enganche” cognitivo corresponden a regiones que participan en la preparación para el movimiento: la corteza prefrontal dorsolateral, los campos oculares frontales, las áreas motoras suplementarias y el lóbulo parietal inferior. Ambas redes, la de enganche cognitivo y la red por defecto muestran oscilaciones en su actividad con una periodicidad de decenas de segundos, que son altamente sincrónicas dentro de cada red, pero notablemente ambas redes son fuertemente antisincrónicas en sujetos normales, inhibiéndose una cuando se activa la otra en forma periódica (22).
En tareas de desempeño contínuo, que consisten en la mantención de respuestas constantes a lo largo del tiempo frente a un estímulo determinado, se ha visto que la variabilidad en los tiempos de respuesta a los estímulos se asocia a la variabilidad en las fluctuaciones de ambas redes (dorsal y red por defecto); es decir, los mayores tiempos de reacción coinciden con los momentos de máxima activación de la red por defecto; y las reacciones más rápidas corresponden a los momentos de máxima activación de la red de enganche cognitivo (22). Siguiendo esta evidencia, Sonuga-Barke y Castellanos (23) propusieron que en el TDAH (y en otras condiciones neuropsiquiátricas), la red por defecto mantendría un estado activado e interferiría con la consolidación de las redes de enganche cognitivo, produciendo un desempeño cognitivo subóptimo. Esta propuesta se apoya en la gran variabilidad en los tiempos de reacción durante tareas de desempeño continuo que se observa en las respuestas de los sujetos con TDAH (24). Sin embargo, hay que notar que aunque algunos estudios sugieren una disminución de la antisincronía entre ambas redes en el TDAH (25), otros estudios han reportado una disminución en la conectividad funcional dentro de la red por defecto en estos pacientes (por ejemplo, 26).
También se ha descrito una tercera red atencional, que comparte características tanto de la red por defecto y de la red de enganche cognitivo (también llamada red dorsal). Esta red, que podría mediar en la reorientación atencional (ya sea desplazando la atención desde un estímulo hacia otro, o desde un período de introspección, comandado por la DMN, a otro de atención focalizada), ha sido descrita como la red ventral (VAN, ventral attention network, 27). Tal como ocurre con la red por defecto, la actividad metabólica de la red ventral disminuye durante tareas con objetivos específicos (como leer o buscar un libro en nuestra biblioteca), pero que se activa transientemente durante la reorientación atencional (por ejemplo cuando se encuentra el libro, o cuando dirijimos rapidamente nuestra atención al escuchar nuestro nombre). Esta red comprende la region supramarginal, el giro temporal superior y la circonvolución frontal media.
Ambas redes (dorsal y ventral) participan en la reorientación atencional, pero desconocemos la manera en la cual interactúan durante este proceso, principalmente por la pobre resolución temporal de la resonancia magnética. Una hipótesis es que el sistema ventral actúa como un corto-circuito que interrumpe la actividad interna (mediada por la red por defecto), modulando la red dorsal cuando aparece un evento relevante o un estímulo inesperado (27). Sin embargo, no está claro dónde comienza la señal de reorientación, ni dónde ocurre la interacción entre estos dos sistemas. Tal cómo reorientar la atención hacia estímulos relevantes es crucial para sobrevivir, reorientarla hacia estímulos irrelevantes puede interferir gravemente en el rendimiento. Por lo tanto, durante cualquier tarea en la cual necesitemos un “enganche” cognitivo, como aprender nueva información durante una clase, se podría generar una ventaja al imponer un filtro atencional, que restrinja la activación ventral (y consecuentemente, la reorientación atencional), protegiendo nuestra atención focalizada de los distractores (28). Por lo mismo, dada la alta resolución temporal de los registros electrofisiológicos, resulta crucial investigar la interacción de dichas redes con esta tecnología, haciendo especial énfasis en la dinámica temporal fina de los procesos mencionados (para la cual los métodos de imaginología son insuficientes), especialmente en trastornos como el TDAH.
Estudios electroencefalográficos de la red en reposoLa red por defecto ha sido también evidenciada usando electroencefalografía (EEG), particularmente a tarvés de correlaciones entre la activación de esta red y la amplitud de oscilaciones de baja frecuencia (29, 30). Usando electrocorticograma se ha podido identificar una activación de alta frecuencia (76-200Hz) en las áreas específicamente asociadas a la red por defecto durante el reposo (31). De la misma manera, usando registros intracorticales, Ossandón y cols. (32) encontraron una supresión de la actividad gama (60-140Hz) en las regiones asociadas a la red por defecto, cuando los sujetos se involucraban en una tarea de búsqueda visual. En TDAH existe evidencia de una disminución de la actividad EEG a bajas frecuencias en la región medial (33), así como de supresión incompleta de la red por defecto durante el enganche cognitivo (34).
También existe evidencia indirecta, proveniente de estudios con potenciales evocados (PE), que apoya la idea acerca de una intromisión de la red por defecto durante la ejecución de tareas cognitivas en TDAH. El potencial P300 es una deflexión positiva de voltaje, localizado en regiones parietofrontales, que se genera aproximadamente 300ms. después de la presentación de un estímulo que requiere de atención o que es relevante para una conducta. Dicho potencial ha sido asociado a la activación de extensas redes de memoria de trabajo durante la ejecución de una tarea, y a otros procesos que revelan el despliegue de diversas funciones cognitivas. Normalmente, el P300 se observa disminuído en ensayos que preceden a respuestas erróneas y en aquellos donde los sujetos reportan haber estado distraídos, tanto niños como adultos (35). En diversas condiciones neuropsiquiátricas, incluído el TDAH, se observa una disminución significativa en la amplitud del P300 en comparación con sujetos normales, que se restablece parcialmente después de medicación (estimulantes, en el caso del TDAH), en concomitancia con un mejor desempeño en las tareas (36).
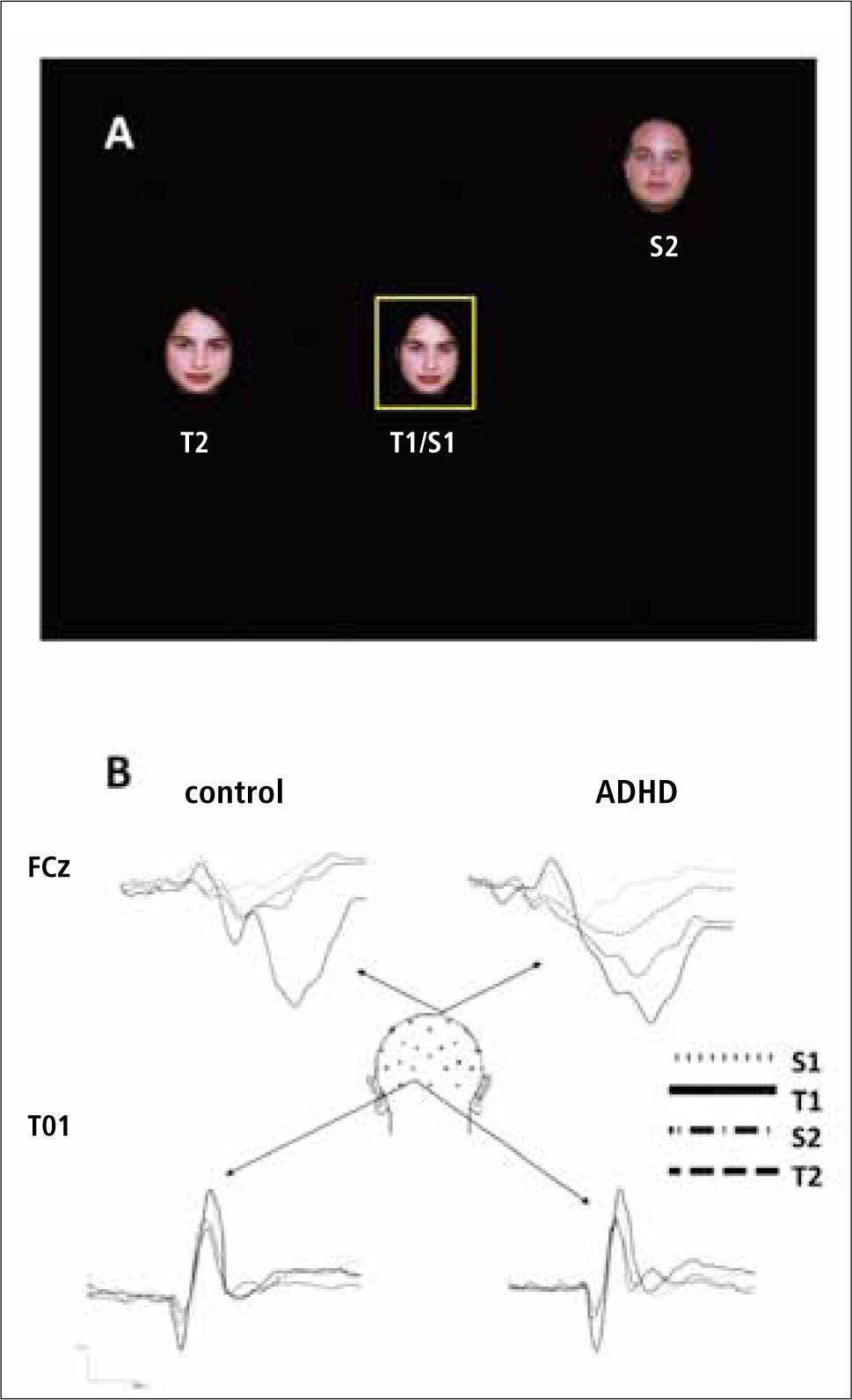
Recientemente, diseñamos dos tareas usando PE para medir la respuesta a estímulos periféricos no atendidos en niños con TDAH, una en el domino espacial y otra en el dominio temporal. La primera de éstas involucraba reconocimiento de caras en el centro de una pantalla donde se presentaban dos caras de distintas personas (una de estas caras dse presentaba con mucha mayor frecuencia que la otra; en relación aproximada 90-10%) (ver Fig. 2). El sujeto debía responder apretando una tecla sólo frente a la presencia de la cara infrecuente (estímulo diana), e ignorar la aparición de la cara frecuente. En estas condiciones, la cara infrecuente (diana) normalmente evoca un robusto potencial P300, en tanto que el estímulo infrecuente no lo hace. Haciendo una variación de esta tarea, mientras los sujetos realizaban la tarea al centro de la pantalla, presentamos también los mismos estímulos en la periferia de la pantalla, pero los sujetos eran explícitamente instruídos para no prestarles atención (37). Tal como se esperaba, los sujetos normales no evidenciaron P300 alguno frente a los estímulos periféricos que no debían ser atendidos, indicando que ellos eran eficaces en suprimir el procesamiento de ellos. Sin embargo, los sujetos TDAH generaron una clara respuesta P300 frente a dichos estímulos periféricos. Esto indica que el estímulo distractor periférico logró desencadenar una forma de actividad cognitiva en los sujetos TDAH, lo cual es consistente con la alta sensibilidad a eventos distractores que caracteriza a la red por defecto.
. En los controles, se observa este potencial (deflexión hacia abajo) sólo con el estímulo T1, que es el que debía ser atendido por el sujeto. Sin embargo, en el grupo TDAH (ADHD en inglés) se ve claramente que los estímulos periféricos, S2 y T2, producen también un P300 significativo, indicando que éstos sujetos no logran suprimir adecuadamente a los estímulos distractores. Los potenciales señalados mas abajo (electrodo T01) son potenciales sensoriales evocados en la corteza visual. Para mas detalles, ver (37).")
B. Potenciales evocados generados durante la tarea. Arriba, el potencial P300 (electrodo FCz). En los controles, se observa este potencial (deflexión hacia abajo) sólo con el estímulo T1, que es el que debía ser atendido por el sujeto. Sin embargo, en el grupo TDAH (ADHD en inglés) se ve claramente que los estímulos periféricos, S2 y T2, producen también un P300 significativo, indicando que éstos sujetos no logran suprimir adecuadamente a los estímulos distractores. Los potenciales señalados mas abajo (electrodo T01) son potenciales sensoriales evocados en la corteza visual. Para mas detalles, ver (37).
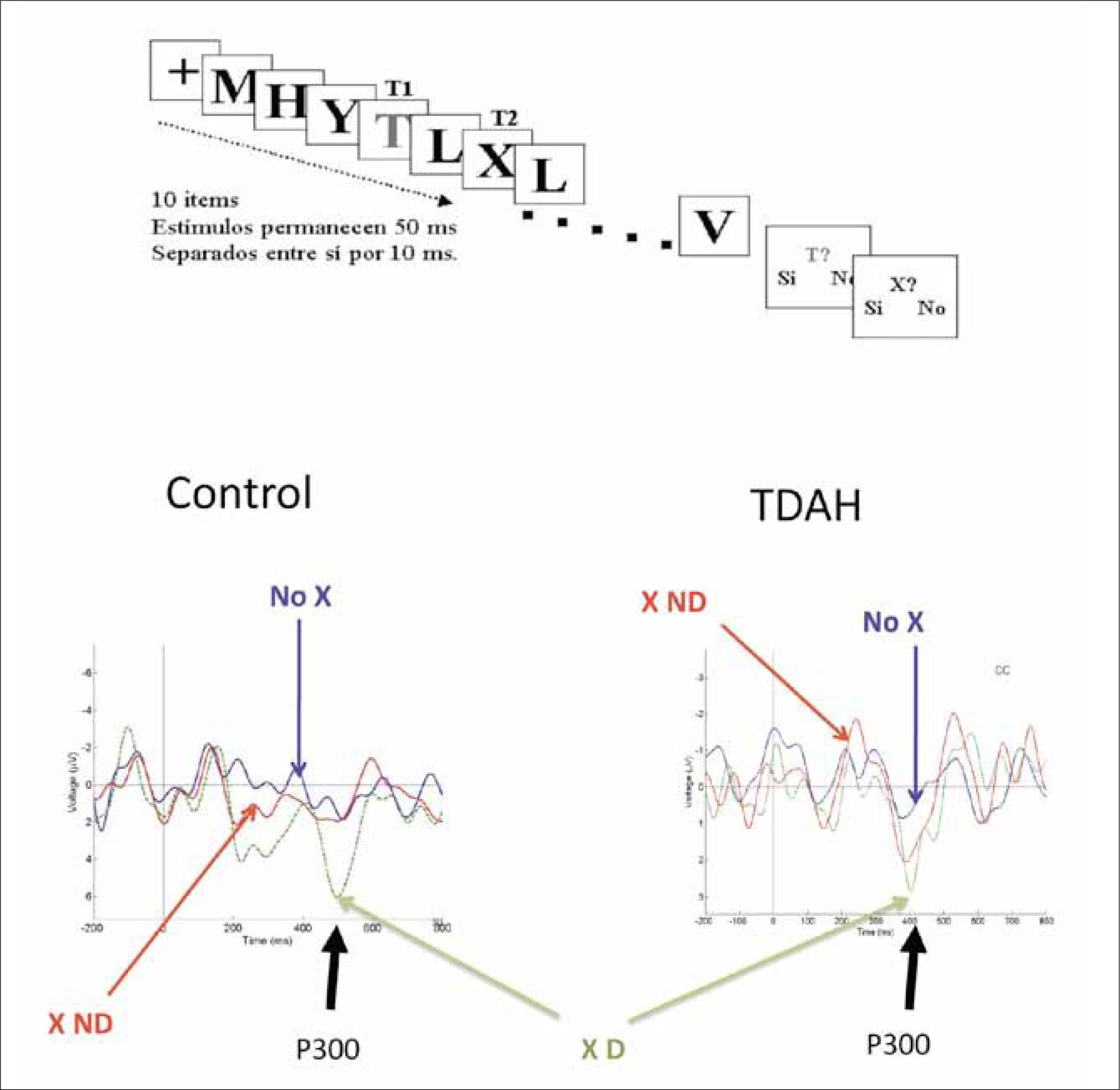
En el segundo experimento, usamos un paradigma de presentación serial visual rápida donde se presenta una serie de imágenes (letras en este caso) muy seguidas en el tiempo (una letra nueva cada 30ms.) y hay que intentar reconocer dos de estas letras en la serie (por ejemplo, reconocer las letras “T” y “X”) (ver Fig. 3). Normalmente, la detección del segundo estímulo es interferida por el reconocimiento del primer estímulo si ambos están demasiado cercanos en el tiempo, fenómeno denominado “parpadeo atencional” (38). Aquí, los controles evidenciaron un robusto P300 frente al segundo estímulo cuando era detectado conscientemente, pero no hubo actividad cuando el estímulo no era percibido, pero estaba presente. Este hallazgo, ya conocido, ha dado lugar a la interpretación de que el P300 es un marcador de la entrada del estímulo a la conciencia. Sin embargo, y en cierta medida contraviniendo esta interpretación, los sujetos TDAH generaron un P300 significativo (aunque de menor amplitud que los controles), tanto cuando percibían el estímulo como cuando no lo percibían, indicando que dicho estímulo era capaz de activar redes que eran normalmente suprimidas en los sujetos control. En ambos paradigmas experimentales, de atención espacial y temporal, encontramos que si bien el potencial P300 era de menor amplitud en los pacientes que en los controles, en el primer grupo también existía un menor umbral para generar este mismo potencial. Nuestra interpretación de estos estudios es que el foco atencional no logra “cerrarse” apropiadamente en estos pacientes, posiblemente debido a la constante intromisión de perturbaciones generadas por la red por defecto.
 una secuencia de 10 letras en la pantalla, entre las cuales hay una letra “T”, de color distinto a las demás, que puede o no ser seguida por una “X”, del mismo color que las otras. Si se detecta correctamente la primera letra, “T”, se hace más difícil poder detectar la segunda letra, “X”, especialmente si ésta es presentada muy cerca de la primera (una o dos letras después).")
A. Diseño experimental en la tarea de parpadeo atencional. Se presenta muy rápidamente (en un intervalo de 600 milisegundos) una secuencia de 10 letras en la pantalla, entre las cuales hay una letra “T”, de color distinto a las demás, que puede o no ser seguida por una “X”, del mismo color que las otras. Si se detecta correctamente la primera letra, “T”, se hace más difícil poder detectar la segunda letra, “X”, especialmente si ésta es presentada muy cerca de la primera (una o dos letras después).
En estas circunstancias, ha sido tentador para muchos autores establecer una asociación entre los mecanismos fásico y tónico de liberación dopaminérgica (o noradrenérgica), con la dinámica de las redes de enganche y por defecto observadas a nivel de activación cerebral (7). En general, tanto la liberación fásica de catecolaminas, como la activación de la red de enganche cognitivo, han sido relacionadas a la conducta orientada a metas, en tanto que niveles bajos de señalización tónica y la red por defecto pueden ser relacionadas con estados de reposo, ensoñación y distractibilidad. Existen varios estudios que han evidenciado una asociación entre la señalización dopaminérgica y las redes de enganche y por defecto en pacientes adultos con la enfermedad de Parkinson y en sujetos adultos normales, que son consistentes con esta hipótesis (39, 40). En esta misma línea, un estudio muy reciente ha demostrado que, en condiciones de reposo, la señalización dopaminérgica facilita el acoplamiento entre la red frontoparietal (VAN) y la red por defecto, a la vez que reduce el acoplamiento de la primera con la red dorsal de enganche cognitivo (41).
A nuestro juicio, los mecanismos moleculares que subyacen a la dinámica de las redes involucradas en los diversos estados cerebrales es una materia de una inmensa proyección tanto básica como clínica. Como vemos, los polimorfismos genéticos pueden generar sesgos en dichas dinámicas, poniendo a los sujetos ya sea más o menos en riesgo de un desarrollo anormal de dichas redes, en condiciones ambientales determinadas. El caso del TDAH se presenta hoy en día como un interesante modelo genético-neuro-cognitivo para el análisis multidimensional de estos fenómenos, dada su prevalencia, su alta heredabilidad, y principalmente el hecho que se trata de una condición que no es invalidante sino que refleja en parte una porción del espectro normal de conductas. Esperamos continuar en esta línea de investigación de manera de aportar en este conocimiento, y de contribuír a que dichos pacientes eventualmente logren acceder a una mejor calidad de vida. Por último, a pesar de estos avances en la neurobiología del TDAH, debemos aclarar que existen elementos ambientales que pueden afectar en forma crítica el desarrollo de ésta condición. El ambiente familiar y el contexto cultural pueden ser factores gravitantes en la sintomatología; de la misma manera, la experiencia subjetiva de estos niños es una dimensión que posiblemente es mejor abordada desde una perspectiva psicoterapéutica (42). Así, el establecimiento de normas conductuales y las conductas de contensión emocional hacia dichos pacientes son elementos fundamentales en el buen manejo de ellos (43).
Los autores declaran no tener conflictos de interés, con relación a este artículo.