





Entre los principales componentes de la cerveza, los alcoholes superiores son de importancia por su influencia en el sabor y el aroma del producto final, y por ende, en su calidad. Durante el proceso de producción de cerveza, estos compuestos se generan por las levaduras a partir del metabolismo de aminoácidos. Tanto las levaduras como las condiciones de fermentación y la composición del mosto afectan el perfil de alcoholes superiores y sus concentraciones. En la presente revisión se reúne información detallada sobre las enzimas que participan en la formación de alcoholes superiores y su regulación. Además, se describe cómo el tipo de levadura utilizada, la temperatura de fermentación, la composición de hidratos de carbono y la fuente de nitrógeno presente en el mosto, entre otros parámetros de fermentación, afectan la biosíntesis de estos alcoholes. El análisis de los factores que afectan los niveles de alcoholes superiores durante la elaboración de la cerveza brinda a los productores cerveceros una herramienta de relevancia para lograr las características deseadas en el producto final, y al mismo tiempo, pone en evidencia aspectos aún desconocidos para la ciencia.
Among the main beer components, fusel alcohols are important because of their influence on the flavor of the final product, and therefore on its quality. During the production process, these compounds are generated by yeasts through the metabolism of amino acids. The yeasts, fermentation conditions and wort composition affect fusel alcohols profiles and their concentrations. In this review, we provide detailed information about the enzymes involved in fusel alcohols formation and their regulation. Moreover, we describe how the type of yeast used, the fermentation temperature and the composition of carbohydrates and nitrogen source in wort, among other fermentation parameters, affect the biosynthesis of these alcohols. Knowing how fusel alcohol levels vary during beer production provides a relevant tool for brewers to achieve the desired characteristics in the final product and at the same time highlights the aspects still unknown to science.
La cerveza es una mezcla compleja, no solo por los compuestos que aportan las materias primas utilizadas para su elaboración (agua, malta, lúpulo y levadura), sino porque durante las diferentes etapas del proceso se producen entre ellos numerosas reacciones e interacciones que definen las características organolépticas (o el «flavor») de la cerveza38.
Las levaduras son el ingrediente más activo en términos generales respecto de su influencia en el flavor, ya que pueden producir numerosos compuestos que inciden tanto de manera positiva como negativa en la calidad de la cerveza. Entre ellos se destacan los ésteres, los alcoholes superiores, los fenoles, los compuestos sulfurados, los aldehídos, las cetonas y los ácidos orgánicos, entre otros. En una publicación anterior se abordó el tema de la síntesis y la regulación de los ésteres por parte de las levaduras cerveceras56; en el presente trabajo nos enfocamos, particularmente, en los alcoholes superiores.
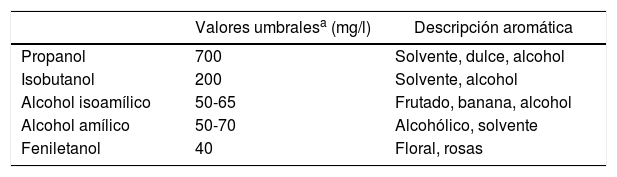
Los alcoholes superiores son el producto del metabolismo secundario de las levaduras, se generan en altas concentraciones respecto de otros compuestos volátiles y son de gran interés por su impacto en el aroma y el sabor de las cervezas68. Este tipo de alcoholes poseen más de dos carbonos y su peso molecular y punto de ebullición son mayores que los del etanol. En cerveza se pueden encontrar aproximadamente 40 alcoholes superiores diferentes58; los que más influyen en el flavor de las cervezas son el propanol, el isobutanol, el feniletanol, el alcohol amílico y el alcohol isoamílico (tabla 1). Este último es el que se encuentra en mayor proporción y el que más afecta la tomabilidad de la cerveza, dado que a concentraciones elevadas genera un intenso flavor a solvente63.
Alcoholes superiores más relevantes en el flavor de la cerveza
| Valores umbralesa (mg/l) | Descripción aromática | |
|---|---|---|
| Propanol | 700 | Solvente, dulce, alcohol |
| Isobutanol | 200 | Solvente, alcohol |
| Alcohol isoamílico | 50-65 | Frutado, banana, alcohol |
| Alcohol amílico | 50-70 | Alcohólico, solvente |
| Feniletanol | 40 | Floral, rosas |
Al igual que la mayoría de los compuestos del flavor, los alcoholes superiores pueden tener un efecto positivo o negativo según su concentración. Sin embargo, en este caso es más probable que su aporte sea negativo, por lo que la comprensión de los factores que afectan su síntesis es muy importante. Altas concentraciones de alcoholes superiores (>300mg/l) en cerveza generan un flavor fuerte (a solvente) y pungente, lo que crea una sensación de calentamiento en boca, mientras que niveles adecuados pueden incrementar, en ciertos estilos, la complejidad de bebidas fermentadas intensificando la percepción alcohólica38,63. Asimismo, la relación entre la cantidad de alcoholes superiores y otros compuestos volátiles es de gran importancia65,71. En cervezas Lager, es deseable una relación 4:1 o 3:1 de alcoholes vs. ésteres. Un aumento en esta relación, provocada por un incremento de alcoholes superiores, da como resultado cervezas más secas y con aromas de menos carácter2.
Los alcoholes superiores son generados por las levaduras durante la fase lag (o de aclimatación) de la fermentación, a partir de la síntesis o la degradación de aminoácidos98 (utilizando piruvato y acetil-CoA en el primer caso o después de la asimilación de la fuente de nitrógeno en el segundo). Es decir que la acumulación de alcoholes superiores en la cerveza es el resultado de la reproducción activa de la levadura, que demanda de aminoácidos para fabricar proteínas. El catabolismo de aminoácidos, después de que estos ingresan a las células de levadura, puede llevarse a cabo a través de la vía de Ehrlich. Esta vía incluye reacciones de transaminación, descarboxilación y reducción para dar lugar a los alcoholes correspondientes, los cuales posteriormente son excretados al medio34.
La producción de alcoholes superiores varía con las cepas de levadura utilizada20. No obstante, otros parámetros fermentativos como la temperatura de fermentación, la aireación y el contenido de nitrógeno en el mosto también influyen en los niveles producidos de estos compuestos45,47,48. En el presente trabajo se describe el modo en que las levaduras sintetizan alcoholes superiores, así como también las enzimas involucradas en este proceso y su regulación. Además, se analiza cómo diferentes factores relevantes para producción de cerveza afectan las concentraciones de este tipo de compuestos del flavor.
Es innegable el crecimiento del segmento de cervezas artesanales: se anticipa un incremento anual a nivel mundial del 9,3% en el período 2015-20207. Para lograr un producto de calidad y acorde con la demanda de los consumidores, los productores de este tipo de cervezas —a menudo insertos en mercados incipientes, aunque promisorios, como es el caso de la Argentina— demandan cada vez más asesoramiento acerca del proceso de producción en general y sobre el manejo de levaduras y el control del flavor generado en la cerveza, en particular. Se pretende que esta revisión, junto con la anterior de la misma serie56, no solo ayuden a dar cuenta de lo que se conoce hasta el día de hoy respecto de cómo se generan los compuestos del flavor, sino que también se constituyan en una herramienta útil para los productores de cerveza artesanal en su camino por obtener los aromas y sabores deseados en sus productos.
¿Cómo se generan los alcoholes superiores en la cerveza?Los alcoholes superiores son formados por las levaduras, a partir de la incorporación y el metabolismo de los aminoácidos presentes en el mosto (fig. 1). El ingreso de los aminoácidos en las células de levaduras es llevado a cabo por proteínas transportadoras ubicadas en la membrana plasmática, las cuales poseen propiedades diferentes en cuanto a su especificidad, afinidad por sustratos, capacidad y regulación5,39. Los aminoácidos ramificados como la valina, la leucina y la isoleucina, requeridos para la síntesis de isobutanol, alcohol isoamílico y alcohol amílico, respectivamente, pueden ingresar a la célula por medio de una permeasa general (GAP), de las permeasas BAP2 y BAP3, con alta afinidad por estos aminoácidos, y de otras permeasas menos específicas, como TAT1, AGP2 y AGP331,77.
, las flechas en azul, verde y rojo indican la acción de enzimas aminotransferasas, descarboxilasas y deshidrogenasas, respectivamente. El color de esta figura solo puede apreciarse en la versión electrónica del artículo.")
Esquema de las principales rutas metabólicas que contribuyen a la síntesis de alcoholes superiores en levaduras inoculadas en mosto cervecero. En la membrana plasmática, BAP2,3, AGP, TAT y GAP representan proteínas transportadoras de aminoácidos. Dentro de la vía de Ehrlich (citoplasma), las flechas en azul, verde y rojo indican la acción de enzimas aminotransferasas, descarboxilasas y deshidrogenasas, respectivamente. El color de esta figura solo puede apreciarse en la versión electrónica del artículo.
Grauslund et al.31 detectaron por primera vez el gen BAP2 en 1995. En ese mismo trabajo se indica que la deleción del gen BAP2 en una cepa de Saccharomyces cerevisiae cuyo alelo GAP1 también se encuentra delecionado (Δgap1) reduce en un 45%, aproximadamente, la absorción de valina y leucina, y en cerca de un 25% la absorción de isoleucina, respecto de una cepa control que contiene deleciones en el gen GAP1 únicamente. La incorporación de otros aminoácidos hidrofóbicos no se vio afectada por la deleción de BAP231. La transcripción de este gen depende de la fuente de nitrógeno presente en el medio y responde de manera opuesta a la transcripción del gen GAP1. Este último gen se encuentra sujeto a la represión por fuentes de nitrógeno fácilmente asimilables, como amonio, asparagina o glutamina21,78. BAP2 puede expresarse en un medio mínimo que contiene amonio como única fuente de nitrógeno, pero los niveles de transcripto aumentan con el agregado de cantidades micromolares de leucina. Esta inducción se ve acompañada no solo de la incorporación de leucina en la célula, sino también de isoleucina y valina21.
En condiciones típicas de producción industrial de cerveza, la formación de isobutanol, alcohol isoamílico y alcohol amílico se encontró asociada a la absorción de aminoácidos ramificados, particularmente al emplear la levadura Lager S23 durante la fermentación. Asimismo, diferencias en la transcripción de permeasas presentes en S23 y S81 (levaduras Lager y Ale, respectivamente) sugieren que estas enzimas podrían afectar la incorporación de aminoácidos y, por lo tanto, las concentraciones de alcoholes superiores logradas por estas cepas66. Esta hipótesis, sin embargo, aún debe ser testeada a fin de confirmar que existe una relación entre los niveles de estos transcriptos y la producción de metabolitos involucrados en la formación del flavor. La concentración de alcoholes superiores depende también de la expresión y actividad de las enzimas involucradas en su síntesis.
Una vez dentro de la célula, los aminoácidos como isoleucina, leucina, la valina, tirosina, el triptofano, metionina y la fenilalanina pueden ser catabolizados mediante la vía de Ehrlich para dar lugar a la formación de alcoholes superiores, y para que esto ocurra deben actuar enzimas transaminasas, descarboxilasas y alcohol deshidrogenasas26. Estos compuestos también se producen durante la biosíntesis de dichos aminoácidos32, ya que los esqueletos carbonados generados durante este proceso son capaces de entrar en la vía de Ehrlich como oxoácidos. La combinación precisa de enzimas activas en un determinado momento depende del aminoácido presente en el medio, del suplemento de oxoácidos y del estado de crecimiento del cultivo20,76. Asimismo, de acuerdo con las necesidades celulares, las vías catabólicas y biosintéticas pueden relacionarse. Por ejemplo, el aminoácido valina es capaz de generar tanto isobutanol como alcohol isoamílico, dado que el catabolismo de este aminoácido y la síntesis de leucina comparten un pool común de alfa-cetoisovalerato18.
Vía de EhrlichAminotransferasasEn la levadura S. cerevisiae, la última reacción en la biosíntesis de aminoácidos y la primera involucrada en su catabolismo se debe a la acción aminotransferasas. Estas enzimas catalizan la transferencia de grupos amino entre aminoácidos y sus correspondientes alfa-cetoácidos, precursores de los alcoholes superiores. Cuatro aminotransferasas se encuentran implicadas en la primera reacción de la vía de Ehrlich. Estas son codificadas por los genes BAT1 (Twt1 o Eca39), BAT2 (Twt2 o Eca40), ARO8 y ARO934. Las enzimas BAT1 y BAT2 participan de la transaminación de aminoácidos ramificados y sus funciones se ven reflejadas en su localización celular y sus patrones de expresión. BAT1 se encuentra ubicada en la mitocondria, mientras que su isoenzima BAT2 se localiza en el citosol25. Asimismo, la isoenzima mitocondrial presenta una alta expresión durante el crecimiento exponencial en cultivos batch y un carácter predominantemente biosintético, aunque también posee un rol catabólico, evidenciado cuando el gen BAT2 es disfuncional15. Su contraparte citosólica, en cambio, se expresa preferentemente durante la fase estacionaria y se encuentra involucrada casi exclusivamente en el catabolismo de la valina, la isoleucina y la leucina15,34.
Algunos trabajos en los que se estudió el efecto de la deleción de estos genes mostraron que la ausencia de ambos conduce a un retardo en el crecimiento y una menor producción de alcoholes superiores15,76. Asimismo, y a pesar de que la mutación simple del gen BAT1 no disminuye de manera significativa la producción de alcoholes superiores, se observaron efectos drásticos en la producción de isobutanol, alcohol amílico e alcohol isoamílico tras la deleción del gen BAT224,76,99. Más aún, la sobreexpresión de este gen puede aumentar hasta dos veces la producción de isobutanol14. Estos resultados indicaron que la expresión de BAT2 sería esencial para la formación de estos compuestos, principalmente durante el crecimiento de las levaduras en etanol76. No obstante, otras transaminasas podrían estar involucradas en la formación de estos alcoholes24,99.
ARO8 y ARO9 son dos aminotransferasas que poseen una amplia especificidad de sustrato y, en la vía de Ehrlich, catalizan la transaminación de aminoácidos aromáticos89. Ambas enzimas difieren en su perfil de expresión. Al igual que BAT1, ARO8 presenta una mayor expresión al inicio de la fermentación respecto de ARO966. ARO8 también se expresa constitutivamente bajo el control de la síntesis de aminoácidos, mientras que la expresión de ARO9 es inducida en presencia de aminoácidos aromáticos40. Boer et al.8 arribaron a una conclusión similar al observar una regulación positiva en la transcripción de ARO9 y BAT2 cuando a cultivos de S. cerevisiae se agregó fenilalanina, metionina y leucina como únicas fuentes de nitrógeno.
Recientemente, a partir del perfil global de transcripción en levaduras Ale y Lager, se analizó la relación entre la expresión de genes asociados a la formación del flavor y los niveles de compuestos volátiles66. La transcripción de los genes BAT1 y ARO8 fue menor en la levadura de tipo Lager, lo cual podría explicar los menores niveles generados de isobutanol, alcohol amílico e alcohol isoamílico en comparación con la levadura de tipo Ale66. No se han encontrado estudios en los que la sobreexpresión de ARO8 y ARO9 se vincule con aumentos de los niveles de feniletanol.
DescarboxilasasLa descarboxilación de los alfa-cetoácidos derivados de la transaminación de los aminoácidos representa la única reacción irreversible en la vía de Ehrlich. Esta etapa es catalizada por descarboxilasas dependientes de tiamina pirofosfato. S. cerevisiae posee cinco descarboxilasas de este tipo, PDC1, PDC5, PDC6, ARO10 y THI3, las cuales conforman una familia de proteínas estrechamente relacionadas20. Los primeros estudios acerca de estas enzimas mostraron que en la descarboxilación de cada oxoácido proveniente del catabolismo de la valina, la leucina y la isoleucina participaba una descarboxilasa diferente, o un grupo determinado de ellas, lo que indica una alta especificidad en esta etapa de la vía de Ehrlich18,19. No obstante, en ese entonces se desconocía la descarboxilasa codificada por el gen ARO10 y, hasta el momento, no hay pruebas de que el gen THI3 realmente codifique una descarboxilasa con actividad catalítica19,72,93.
Una caracterización más profunda de estas enzimas utilizando una combinación de estrategias fisiológicas, genéticas y bioquímicas demostró luego que las tres isoenzimas PDC pueden descarboxilar sustratos aromáticos como el fenilpiruvato y los 2-oxoácidos derivados de aminoácidos ramificados y azufrados, sin diferir de manera significativa en su especificidad, a excepción de PDC5, que posee mayor actividad frente a fenilpiruvato y una Km más alta para el piruvato72,93. De las cinco descarboxilasas, PDC1, PDC5 y PDC6 son las únicas capaces de catalizar la descarboxilación de 2-oxoácidos lineales como el 2-cetobutanoato y el 2-cetopentanoato, precursores de n-propanol y n-butanol, respectivamente72.
Las isoenzimas PDC, por otra parte, se expresan de manera diferencial. La enzima PDC1 es capaz de expresarse en casi cualquier condición, mientras que la PDC5 presenta una mayor transcripción en ausencia del gen PDC176,79, bajo limitación de nitrógeno8,87 y en cultivos limitados en tiamina61. Los niveles de expresión de PDC6 son mayores en cultivos limitados en azufre87.
La ARO10 es una descarboxilasa con amplia especificidad de sustrato, cuya actividad depende de la fuente de nitrógeno utilizada durante el crecimiento de las levaduras. Extractos celulares de una cepa de S. cerevisiae carente de los genes PDC1, PDC5, PDC6, THI3 y ARO10, a la que se incorporó un vector con el gen ARO10 bajo el control de un promotor constitutivo, mostraron actividad descarboxilasa sobre 2-oxoácidos derivados de la transaminación de la valina, la isoleucina, la leucina, la metionina y la fenilalanina72,92.
En comparación con las tres enzimas PDC, ARO10 presentó mayor afinidad por estos sustratos, de modo que esta podría ser la principal descarboxilasa responsable de la formación de alcoholes superiores relevantes en el flavor de cervezas72,91. La transcripción de ARO10 es inducida por leucina, metionina, triptofano y fenilalanina, aunque es mayor en cultivos que contienen aminoácidos aromáticos como única fuente de nitrógeno8,72,92,93. La regulación transcripcional de ARO10 es mediada por el regulador positivo ARO80, cuya síntesis se encuentra sometida a la represión por catabolitos nitrogenados (NCR). Este regulador se une constitutivamente a promotores de ARO9 y ARO10 favoreciendo así la expresión de estos genes en cultivos carentes de fuentes de nitrógeno fácilmente asimilables. Además, los aminoácidos aromáticos activan el regulador ARO80 e incrementan aún más la transcripción de ARO9 y ARO10, ejerciendo así un efecto aditivo en la regulación de ambos genes46. Por otro lado, Vuralhan et al.92,93 observaron que los niveles de transcripto no se correlacionan con la actividad descarboxilasa, lo cual sugiere que la regulación de la expresión de ARO10 podría ocurrir en la etapa postranscripcional.
Diferentes alelos ARO10 pertenecientes a distintas levaduras también pueden codificar enzimas ARO10 con una amplia especificidad de sustrato, aunque difieren en su regulación y actividad9,83. En la levadura cervecera Saccharomyces pastorianus CBS1483 (híbrido de S. cerevisiae x Saccharomyces eubayanus), si bien la expresión de los alelos ARO10 correspondientes a cada subgenoma (LgSeubARO10 y LgScARO10) se induce en presencia de fenilalanina, dicha inducción es mayor en el alelo LgScARO10. Más aún, los niveles de transcripto de LgSeubARO10 se ven incrementados en presencia de fuentes de nitrógeno como amonio y leucina, no así su contraparte derivada del subgenoma de S. cerevisiae (LgScARO10)9. Ante estos resultados, Bolat et al.9 sugirieron que LgSeubARO10 y LgScARO10 cumplirían diferentes roles. Por su expresión constitutiva en ausencia de aminoácidos extracelulares, LgSeubARO10 estaría involucrada en la producción de alcoholes superiores a partir de la síntesis de aminoácidos, mientras que LgScARO10 contribuiría a su formación mediante la vía de Ehrlich. Además, a pesar de que ambos alelos mostraron una actividad similar frente a la mayoría de los sustratos evaluados, con preferencia por el fenilpiruvato, la actividad de la descarboxilasa codificada por LgSeubARO10 frente a ceto-isovalerato (compuesto que forma parte de la síntesis de la leucina y del catabolismo de la valina) fue dos veces mayor. En contraposición con las descaboxilasas anteriores, Strybny et al.83 observaron que la enzima SkARO10 de Saccharomyces kudriavzevii (especie capaz de hibridar con S. cerevisiae para dar lugar a levaduras utilizadas en la producción de cervezas belgas) posee una actividad similar con cada sustrato evaluado (derivados de valina, leucina, isoleucina, fenilalanina y metionina), sin preferencias por ninguno en particular. No obstante, fermentaciones en mosto sintético realizadas con una cepa de S. cerevisiae a la que se le cambió el gen ScARO10 por SkARO10 dieron lugar a mayores concentraciones de alcohol isoamílico e isobutanol respecto de la cepa que contenía ScARO10. Estas diferencias podrían deberse a la complejidad del medio y al impacto de otros compuestos en la formación de alcoholes superiores83.
A partir de la selección a gran escala de genes de S. cerevisiae cuya deleción afecta la producción de compuestos aromáticos, se identificaron genes codificantes de otras dos descarboxilasas (PAD1 y SPE1) que podrían ser de importancia en la vía de Ehrlich84,85. Ambas enzimas tendrían la capacidad de actuar de manera promiscua y, por lo tanto, aceptar varios compuestos como sustratos, como por ejemplo, los ácidos p-cumárico y ferúlico. Ambos ácidos son precursores para la formación de fenoles, compuestos que también influyen en el flavor de las cervezas11. Sin embargo, se necesitan más estudios bioquímicos para confirmar las propiedades de estas enzimas y su rol directo en el catabolismo de aminoácidos84.
DeshidrogenasasLa última etapa de la vía de Ehrlich consiste en la reducción del aldehído formado durante la descarboxilación, reacción catalizada por enzimas alcohol deshidrogenasas. El genoma de S. cerevisiae contiene genes codificantes de 16 alcohol deshidrogenasas, 6 aldehído deshidrogenasas y al menos dos reductasas de amplio espectro dependientes de nucleótidos pirimidínicos34. Cualquiera de las enzimas etanol deshidrogenasas ADH1, ADH2, ADH3, ADH4 y ADH5, junto con la formaldehído deshidrogenasa SFA1, pueden tener un papel significativo en la formación de alcoholes superiores a través de la vía de Ehrlich20, aunque Styger et al.84 observaron que en medio complejo solo la deleción del gen ADH3 afecta la producción de estos compuestos. ADH3 es responsable de la reducción de acetaldehído dentro de la mitocondria, lo que mantiene el balance rédox durante el crecimiento de las levaduras en anaerobiosis4. Los aldehídos involucrados en esta etapa de la vía pueden dar lugar tanto a alcoholes superiores como a ácidos; que predomine una u otra reacción depende del estado rédox de las células84,93.
Otras deshidrogenasas podrían estar directamente implicadas en la vía de Ehrlich. Entre ellas se encuentran OYE2, AAD6 y HOM2. OYE2 es capaz de oxidar tanto NADPH como NADH, y de reducir diferentes sustratos10,88. La deleción del gen codificante de esta enzima provoca una notable disminución en los niveles de isobutanol e alcohol isoamílico84. En S. cerevisiae, el gen AAD6 se transcribe mayormente al inicio de la fermentación y su deleción también lleva a menores concentraciones de estos compuestos66,84. Dickinson et al.20 no observaron tales efectos cuando se delecionaron los 7 genes AAD presentes en S. cerevisiae, lo cual podría deberse a diferencias en el medio de cultivo utilizado84. HOM2 es una semialdehído deshidrogenasa que cataliza el segundo paso de la biosíntesis de la metionina y la treonina3. En comparación con los genes AAD6 y OYE2, HOM produce una mayor reducción en la concentración de alcoholes superiores cuando es eliminado84. Durante el transcurso de la fermentación, la transcripción de HOM2 es regulada positivamente66.
La vía de Ehrlich permite explicar solo la formación de alcoholes como isobutanol, alcohol isoamílico, alcohol amílico y feniletanol, no así la producción de otros alcoholes superiores, dado que no todos se corresponden con los aminoácidos conocidos20. Se cree que una de las razones por las cuales se forman los alcoholes superiores es para mantener la relación NADH/NAD+ y, por lo tanto, el balance redox dentro de la célula22. Sin embargo, hay quienes sostienen que compuestos como el etanol, el glicerol, el acetato, el acetaldehído y el succinato también cumplen este rol. Otra posibilidad es que los alcoholes superiores se generen para remover aldehídos tóxicos para las células, o como fuente alternativa para obtener nitrógeno84,93.
El mecanismo por el cual se liberan estos compuestos al mosto aún se desconoce. Hasta el momento no se han detectado transportadores de membrana que intervengan en la excreción de alcoholes superiores, por lo que es posible que este proceso ocurra simplemente por difusión pasiva a través de la membrana celular de las levaduras52.
Factores que afectan la formación de alcoholes superioresComposición del mostoEl material de partida para la elaboración de cerveza, es decir, la composición del mosto, juega un papel importante en el perfil de flavor del producto final. En general, para la producción de cerveza se utilizan granos de cebada malteada, pero también se pueden incluir otros cereales como trigo, avena, sorgo y centeno38,57. Dependiendo del tipo de grano utilizado, el mosto generado puede tener una composición de azúcares y nitrógeno diferente, lo cual afecta de manera significativa los niveles de alcoholes superiores generados por las levaduras17.
Efecto de la composición de hidratos de carbonoEn la composición de un mosto estándar hay aproximadamente un 90% de hidratos de carbono, que incluyen azúcares como glucosa, fructosa, sacarosa, maltosa, maltotriosa y dextrinas. El perfil de azúcares y su concentración tiene una influencia directa en la eficiencia de fermentación y en el metabolismo de la levadura. Al incrementarse los niveles de sacarosa en el mosto se eleva la producción de alcoholes superiores36, mientras que el metabolismo de la maltosa disminuye la concentración de estos compuestos en comparación con fuentes de carbono más fácilmente asimilables, como glucosa y fructosa48. Es por esto que cuando se agregan adjuntos a fin de aumentar la densidad inicial del mosto, se debe tener en consideración el tipo de azúcar agregado. En un trabajo reciente donde se utilizó S. pastorianus, se observó que el suplemento de glucosa en mostos con densidades de 18 y 24°P incrementó entre un 17% y un 90% las concentraciones de isobutanol y feniletanol respecto de mostos de la misma densidad a los que se agregó sacarosa y maltosa48. En cambio, el agregado de maltosa solo aumentó los niveles de alcohol isoamílico en los mostos analizados48. La adición de este azúcar podría dar origen a un perfil del flavor más favorable en el producto final, dado que a su vez disminuye la formación de ésteres de acetato56,64, lo cual facilitaría el diseño de mostos de alta densidad.
En mostos con 18°P de densidad inicial fermentados con levaduras de tipo Ale se ha observado un incremento en los niveles de alcoholes superiores respecto de mostos de densidad inicial menor73. Que estos compuestos aumenten con la densidad del mosto se correlaciona con el hecho de que altas concentraciones de hidratos de carbono afectan de manera positiva la actividad de enzimas aminotransferasas24,70. No obstante, en levaduras Lager, la producción de estos compuestos puede no verse afectada o disminuir en mostos más concentrados48,49,100. El genoma de levaduras Lager presenta un alto grado de plasticidad: es capaz de experimentar cambios genéticos en respuesta a condiciones de estrés tales como mostos de alta densidad y altas temperaturas41. Por otro lado, en mostos con elevada relación azúcares/nitrógeno, la actividad fisiológica normal de las levaduras se ve modificada, lo cual genera desbalances en la producción de compuestos del flavor36,81. En este caso, la fuente de nitrógeno se convierte en el factor limitante en el metabolismo de las levaduras. De hecho, dado que la producción de alcoholes superiores se encuentra directamente ligada al metabolismo de los aminoácidos, la fuente de nitrógeno presente en el mosto es el principal sustrato que influye en su biosíntesis.
Efecto de la fuente de nitrógenoEn general, un aumento en la concentración de nitrógeno asimilable permite incrementar la producción de compuestos del flavor, como alcoholes superiores y ésteres33,50.
La transcripción de genes asociados a la síntesis de estos compuestos puede verse afectada al aumentar los niveles de la fuente de nitrógeno en el mosto49,73,99. El tipo de aminoácidos que forman parte del nitrógeno asimilable genera diferentes respuestas en las levaduras y también influye en la formación de alcoholes superiores. En medio sintético, el aminoácido leucina en cantidades micromolares actúa como el principal regulador de señal para la incorporación de aminoácidos ramificados en S. cerevisiae21,97. Como se mencionó anteriormente, la incorporación de leucina, isoleucina o valina y su asimilación a través de la vía de Ehrlich provocan un aumento en la cantidad de alcohol isoamílico, alcohol amílico, isobutanol, respectivamente, y sus correspondientes ésteres de acetato34,69,82. Además, el análisis de los perfiles de expresión de S. cerevisiae durante la formación de compuestos del flavor cuando es cultivada en L-leucina y amonio reveló una regulación positiva o negativa en 117 genes, entre los cuales se incluyeron genes codificantes de enzimas involucradas en diferentes rutas del metabolismo de aminoácidos75.
La prolina y la histidina son otros aminoácidos de relevancia para el flavor de las cervezas50,67. Fermentaciones realizadas en medio sintético con adición de prolina utilizando levaduras de tipo Ale y Lager permiten llegar a mayores concentraciones de alcoholes superiores. Este aminoácido no puede ser transformado en un alcohol superior a través de la vía de Ehrlich, por lo que su incidencia en el flavor podría explicarse por la síntesis de glutamato a partir de dicho aminoácido67. La adición de histidina incrementa de manera significativa la producción de alcoholes superiores como isobutanol, alcohol isoamílico y feniletanol50. Lei et al.50, observaron que en levaduras Lager en fase exponencial se activan genes codificantes para permeasas específicas del transporte de aminoácidos (BAP2, LYP1, HIP1, y AGP1) cuando se utilizan mostos de alta densidad suplementados con histidina. A su vez, durante la fermentación en estas mismas condiciones, se reprimen genes involucrados en el mecanismo de represión del catabolismo del nitrógeno6,50.
La temperatura de fermentaciónLa concentración final de alcoholes superiores en cervezas se ve incrementada al utilizar mayores temperaturas de fermentación37,45,74. El aumento en el crecimiento y el metabolismo de las levaduras al aumentar la temperatura de fermentación favorece la absorción y asimilación de aminoácidos del mosto, por lo que se estima cierta correlación entre la expresión de los genes involucrados en este proceso, la temperatura y los niveles de alcoholes superiores en cerveza45. De hecho, Kodama et al.42, al elevar la temperatura de fermentación, lograron la expresión constitutiva del gen codificante de la permeasa BAP2 presente en la levadura cervecera BH-225, y ello dio lugar a una mayor asimilación de valina, leucina e isoleucina. Aunque en ese estudio se detectó un aumento en la producción de alcohol isoamílico, se sugirió que durante la formación de otros alcoholes podrían utilizarse mecanismos diferentes pero relacionados42. Abe et al.1 también confirmaron la dependencia de la temperatura por parte del gen BAP2, al detectar una disminución en los niveles de expresión de dicho gen a bajas temperaturas de fermentación. En relación con los genes codificantes de las aminotransferasas de la vía de Erhlich, temperaturas más altas incrementan los niveles de expresión del gen BAT1, lo que da lugar a una mayor producción de alcoholes superiores73.
Otros factores que afectan la producción de alcoholes superioresLa formación de alcoholes superiores se ve afectada positivamente en presencia de oxígeno y ácidos grasos insaturados. En mostos con suficiente cantidad de nutrientes, ambos factores (al igual que la temperatura) estimulan el crecimiento de las levaduras y, por ende, el metabolismo de aminoácidos54. En respuesta a la presencia de oxígeno varios genes se expresan diferencialmente, entre los cuales se encuentra el gen codificante de la permeasa BAP2. Al aumentar la concentración de oxígeno, los niveles de transcripto de este gen pueden incrementarse en una hora, y favorecer así la incorporación de aminoácidos en las levaduras y la formación de alcoholes superiores90. Sin embargo, y a pesar de que una mayor oxigenación del mosto puede provocar un aumento en la concentración de estos compuestos, este incremento no es tan significativo como lo es el de la expresión de BAP290. Probablemente el oxígeno regule de manera diferente otros genes que pueden también estar involucrados, directa o indirectamente, en la síntesis de estos compuestos.
A diferencia de lo que sucede con el oxígeno, los niveles de alcoholes superiores prácticamente no se ven afectados por el incremento de CO2 durante las primeras etapas de fermentación23. Hacia el final de la fermentación, el aumento en la presión de CO2 puede causar una disminución en la formación de alcoholes superiores, aunque en menor medida respecto de cómo afecta en la síntesis de ésteres56. Esta disminución en la concentración de compuestos del flavor es parcialmente causada por la inhibición del crecimiento de la biomasa metabólicamente activa71.
Las levaduras requieren iones y vitaminas para su eficiente proliferación y fermentación. Estos componentes del mosto permiten que las enzimas y coenzimas de las levaduras funcionen de manera correcta, por lo que una deficiencia de iones o vitaminas puede dar lugar a alteraciones en el metabolismo y afectar el flavor del producto final94. En general, el mosto contiene suficiente cantidad de iones inorgánicos, aunque ocasionalmente podría ser necesario un suplemento adicional (como en el caso del cinc)95. El cinc es de gran importancia en la producción de alcoholes superiores. Cumple funciones catalíticas, estimula la ruptura de alfa-cetoácidos, con lo cual la producción de estos compuestos se ve favorecida23. No obstante, solo concentraciones elevadas de Zn (10ppm) en el mosto se asocian con diferencias significativas en los niveles de alcoholes superiores62.
La tasa de inóculo también puede afectar la producción de alcoholes superiores. Generalmente, el incremento en la concentración de levaduras inoculadas favorece la formación de algunos alcoholes superiores, aunque diferentes autores han aportado resultados contradictorios al respecto. A escala industrial, Kucharczyk et al.44 no observaron un efecto significativo en la cantidad total de alcoholes al modificar la concentración de inóculo inicial de 5x106 a 9x106 células/ml de mosto. Por otro lado, tasas de inoculación demasiado altas pueden disminuir la concentración de alcoholes superiores15, probablemente por una reducción en el crecimiento celular y, por lo tanto, en el metabolismo de los aminoácidos. Al parecer, el impacto de la concentración de inóculo en el flavor depende, además, de la cepa de levadura, su estado fisiológico y las condiciones de propagación23,44,90.
La levadura: factor determinante en el perfil aromático de las cervezasTal como sucede con la producción de ésteres, la cepa de levadura utilizada en la producción de cerveza juega un papel preponderante en la formación de alcoholes superiores56,80.
En general, levaduras de tipo Lager proveen aromas y sabores más limpios, asociados a una producción de bajos niveles de alcoholes superiores y ésteres comparados con levaduras de tipo Ale96. Dentro de las levaduras Lager, a su vez pueden distinguirse dos grupos de levaduras con características bien diferenciadas: grupo I o Saaz y grupo II o Frohberg80. Respecto de la producción de alcoholes superiores, fermentaciones realizadas a temperaturas entre 10 y 14°C con levaduras Lager correspondientes al grupo II son capaces de producir entre 1 y 14mg/l más de isobutanol, alcohol isoamílico y alcohol amílico respecto de aquellas pertenecientes al grupo I28,96. Distintos niveles en la producción de alcoholes superiores entre cepas podrían deberse a diferencias genéticas (por ejemplo, mutaciones puntuales, rearreglos genómicos, deleciones o aneuploidías), regulatorias y de expresión de los genes correspondientes, así como también a diferencias en la especificidad de sustrato de las enzimas codificadas por dichos genes35,43,56.
Particularmente, que las levaduras Lager (S. pastorianus) sean híbridos aneuploides en los que una parte de su genoma proviene de S. cerevisiae y la otra de S. eubayanus (esta última especie, adaptada al frío)51 ha generado interrogantes sobre la transcripción y regulación de los genes involucrados en la producción del flavor en cada subgenoma y su contribución.
Algunos resultados del análisis de la transcripción de los genes BAP2 y BAT2 durante la producción de cerveza con una cepa Lager sugieren una dominancia fenotípica del subgenoma de S. eubayanus respecto de la parte de S. cerevisiae35. En esos trabajos, y en concordancia con lo observado por Kodama et al.42, surgieron indicios de que los genes ortólogos BAP2 difieren en su patrón de expresión en distintas etapas de la fermentación, lo cual indica que en levaduras Lager ambos genes son regulados de manera diferencial y que podrían cumplir diferentes funciones. Esto podría explicar, en parte, la diferencia en el perfil de alcoholes superiores generados por levaduras Lager en comparación con sus parentales. Por otro lado, en la levadura de tipo Lager S. pastorianus CBS1483, Bolat et al.9 describieron roles diferentes para dos isoenzimas descarboxilasas que participan en la síntesis de alcoholes superiores. La especificidad de sustrato de la descarboxilasa codificada por el alelo proveniente de S. cerevisiae fue similar a la observada en una cepa de S. cerevisiae. La misma enzima codificada por el alelo de S. eubayanus, además, presentó mayor preferencia por el compuesto cetoisovalerato, precursor para la síntesis de isobutanol, lo cual podría afectar el balance del flavor respecto de cervezas producidas con cepas de S. cerevisiae9.
Diversos artículos han demostrado el potencial de levaduras no convencionales como herramientas de innovación en la producción de cerveza12,28,60. Brettanomyces bruxellensis, Torulaspora delbruekii, Pichia kluyveri, Naumovia dairenensis y S. eubayanus son algunos ejemplos de estas levaduras, las cuales difieren en su capacidad fermentativa y en la producción de compuestos del flavor, entre otras características27,29. En relación con la producción de alcoholes superiores, levaduras salvajes como S. eubayanus y otras levaduras sacaromicéticas no comerciales tienden a producir altos niveles de estos compuestos29,59. T.delbruekii, en cambio, puede generar menos alcoholes superiores que S. cerevisiae, pero ser apta para la producción de estilos de cerveza como American Amber Ale, Weiss, Stout y Dunkel, dado que brinda niveles agradables de alcohol amílico y aporta complejidad al producto final, lo que da lugar a cervezas de aromas y sabores agradables12,27,60. Por otro lado, el cocultivo de las levaduras salvajes Kazachstania servazzi o N. dairenensis con S. cerevisiae durante la producción de cerveza permitió generar mayores niveles de alcohol isoamílico, feniletanol, feniletil acetato, etil octanoato y etil decanoato, con lo que se obtuvo un flavor más frutado respecto del control30.
Si bien este tipo de levaduras son promisorias a la hora de generar productos novedosos, poco se conoce sobre los genes involucrados en la formación del flavor y su regulación en estas levaduras29. Esto constituye una limitación al momento de ser aplicadas en el sector cervecero, ya que es necesario que el productor sepa cómo controlarlas para poder generar las características deseadas en la cerveza.
Consideraciones finalesDada la gran variedad de compuestos responsables del aroma y sabor en la cerveza, y de los diferentes parámetros que afectan su formación, la mayor parte de la base genética involucrada en la producción de compuestos del flavor y en su variación natural permanece sin conocerse en su totalidad. En el caso de los alcoholes superiores, aún se desconocen enzimas que podrían estar involucradas en la vía de Ehrlich. Una de las dificultades en determinar las enzimas específicas para la producción de alcoholes superiores es el amplio grado de redundancia de los genes correspondientes en el genoma de la levadura38, por lo que la presencia de múltiples genes codificantes de aminotransferasas, descarboxilasas y alcohol deshidrogenasas genera grandes desafíos al momento de estudiar la producción de alcoholes superiores. Por otro lado, el desarrollo de nuevas herramientas para el análisis poligénico ha permitido detectar genes nunca antes asociados a la producción de compuestos del flavor86. A partir del análisis del polimorfismo de nucleótido simple (SNP) en locus de caracteres cuantitativos (QTL) de cepas S. cerevisiae se ha logrado relacionar los genes TOR1 y FAS2 a la producción de fenil etilacetato y alcoholes superiores, como feniletanol, alcohol isoamílico e isobutanol13. El gen TOR1 se encuentra involucrado en la regulación de múltiples procesos celulares, entre los que se encuentran la represión por catabolismo del nitrógeno y la transcripción de aminoácido permeasas16,55. El gen FAS2, en cambio, estaría principalmente ligado al metabolismo del fenil etilacetato y de otros ésteres, dado que codifica la subunidad alfa de la enzima ácido-graso sintasa13.
La concentración de sustratos para las levaduras y la composición del mosto también son de gran importancia en la regulación de los niveles de alcoholes superiores en la cerveza (tabla 2). Si bien en los últimos años se ha avanzado en el conocimiento del impacto de la fuente de carbono y nitrógeno en la regulación de genes de levadura involucrados en la producción de estos compuestos, es necesario realizar más investigaciones al respecto. La asimilación de prolina y su papel en la formación de compuestos aromáticos no se conoce por completo67. Asimismo, dado que la maltosa es el azúcar principal del mosto, sería de gran utilidad conocer qué proporción de diferentes azúcares regula positivamente la expresión de genes responsables de perfiles aromáticos específicos.
Impacto de algunos parámetros fermentativos sobre la producción de alcoholes superiores y la transcripción de genes involucrados en su síntesis
| Concentración de alcoholes superiores | Transcripción de genes | |
|---|---|---|
| ↑ Fuente de nitrógeno | ↑ | ↑ o ↓ Permeasas (GAP1, BAP2)a↑ Aminotransferasa ARO9b↑ Descarboxilasa ARO10b |
| ↑ Fuente de carbono | ↑c | ↑ Aminotransferasasd |
| ↑ Oxígeno | ↑ | ↑ Permeasa BAP2 |
| ↑ Temperatura | ↑ | ↑ Permeasa BAP2↑ Aminotransferasa BAT1 |
| Levadura de tipo Ale (vs. Lager) | ↑ | ↑ Aminotransferasas (BAT1 y ARO08) |
Existen diferentes opciones para el control de la síntesis de alcoholes superiores durante la fermentación (tabla 2). Junto con la combinación adecuada de factores, tales como la temperatura de fermentación y la composición de sustratos, la cepa de levadura es fundamental en la formación de compuestos del flavor. La síntesis de alcoholes superiores puede estar muy bien balanceada si se elige la cepa adecuada53,68.
FinanciaciónLos autores son miembros del CONICET y este trabajo fue financiado por el proyecto PICT 3677 del FONCyT, el proyecto PIP424 del CONICET y la Universidad Nacional del Comahue.
Conflicto de interesesLos autores declaran no tener ningún conflicto de intereses.
Nuestro agradecimiento a los apasionados productores de cerveza artesanal de la Argentina y en particular de la Patagonia quienes inspiraron esta serie de artículos de revisión.
