




La variancia genética del trastorno por déficit de atención e hiperactividad (TDAH) es determinante para el fenotipo. La repetición en tándem en número variable (VNTR) de 40 pares de bases (pb) en la región no traducida 3’ (UTR) del gen DAT1 se ha asociado ala susceptibilidad de presentar TDAH debido al incremento de expresión del transportador de dopamina.
ObjetivoDeterminar la asociación entre el VNTR del DAT1 y el fenotipo y/o endofenotipos del TDAH en una muestra de niños de 6 a 15 años de la ciudad de Bogotá.
Sujetos y métodosSe seleccionó a 73 pacientes con TDAH y 54 controles. En todos los individuos se realizó una prueba de WISC y se valoraron las funciones ejecutivas. Mediante reacción en cadena de la polimerasa se amplificó el VNTR de DAT1. Se establecieron estadísticos genético-poblacionales, análisis de asociación y correlación entre las pruebas neuropsicológicas y el genotipo.
ResultadosEl polimorfismo del DAT1 no mostró asociación con TDAH (p=0,85). Sin embargo, el genotipo 10/10 evidenció asociación con el índice de velocidad de procesamiento (p < 0,05). En el subtipo hiperactividad hubo correlación genotípica con subpruebas de la función ejecutiva (flexibilidad cognitiva) (p ≤ 0,01). En el subgrupo mixto, el genotipo 10/10 se asoció al índice de comprensión verbal del WISC (p < 0,05).
ConclusionesSe encontró una correlación entre el genotipo del VNTR de DAT1 con la subprueba «índice de velocidad de procesamiento» del WISC y la subprueba «flexibilidad cognitiva» de la función ejecutiva. Este es el primer reporte que evalúa el gen DAT1 en población colombiana con TDAH.
Genetic variance of attention deficit-hyperactivity disorder (ADHD) is a strong determinant of this disorder. The 40 base pairs (bp) variable number tandem repeat (VNTR) located in the 3’ untranslated region (UTR) of DAT1 gene increases the expression of the dopamine transporter. Therefore, DAT1 has been associated with susceptibility to ADHD.
ObjectiveTo determine the association between the VNTR of DAT1 and the phenotype of ADHD or its endophenotypes in a sample of children aged between 6 and 15 years from Bogotá.
Subjects and methodsWe selected 73 patients with ADHD and 54 controls. WISC test was applied in all subjects and executive functions were assessed. The VNTR of DAT1 was polymerase chain reaction-amplified. Data regarding population genetics and statistical analysis were obtained. Correlation and association tests between genotype and neuropsychological testing were performed.
ResultsThe DAT1 polymorphism was not associated with ADHD (P=.85). Nevertheless, the 10/10 genotype was found to be correlated with the processing speed index (P<.05). In the hyperactivity subtype, there was a genotypic correlation with some subtests of executive function (cognitive flexibility) (P≤.01). In the combined subtype, the 10/10 genotype was associated with verbal comprehension index of WISC (P<.05).
ConclusionsA correlation was found between DAT1 VNTR and the subtest “processing speed index” of WISC and the subtest “cognitive flexibility” of executive functions. To our knowledge, this is the first report to assess DAT1 gene in a Colombian population.
El trastorno por déficit de atención e hiperactividad (TDAH) es uno de los problemas neuropsiquiátricos más frecuentes en la edad escolar, con una prevalencia mundial que oscila entre el 4 y el 19%1,2. Este trastorno ocupa gran parte de la consulta de psiquiatría infantil y neurología en centros especializados, afectando principalmente al género masculino con una proporción 3:1. Además, produce alteraciones importantes en el rendimiento académico del individuo, en su forma de relacionarse con sus pares y en su vida futura debido a la persistencia de algunos síntomas hasta la edad adulta.
Se han descrito varios factores contribuyentes en la etiología del trastorno, entre los cuales se destacan factores de tipo ambiental, psicológico y genético, por lo que se considera una entidad de origen multifactorial. La heredabilidad estimada del 76% indica que la variancia genética ejerce una alta contribución al desarrollo del fenotipo, por lo que se han planteado varios estudios en la búsqueda de genes candidatos3–5. En concordancia con el mecanismo de acción de algunos fármacos utilizados para mejorar la sintomatología de este trastorno, se han postulado genes involucrados, por un lado en las vías relacionadas con los sistemas dopaminérgico, acetilcolinérgico y serotoninérgico, y por el otro en aquellos que codifican para proteínas transmembranales. Se ha demostrado que los genes que codifican para: receptores del trasportador de dopamina (DAT1), receptores de dopamina (DRD2, DRD3, DRD4 y DRD5), el transportador de serotonina (5HTT), los receptores 1B y 2A para serotonina, la enzima catecol-o-metil transferasa (COMT), la enzima monoamino oxidasa (MAO) y la proteína asociada a sinaptosoma 25 (proteína SNAP25) pueden relacionarse con una mayor susceptibilidad al trastorno4. De hecho, en una revisión de la literatura sobre la genética del TDAH, Faraone et al- examinaron las OR de varios estudios caso-control y estudios de transmisión alélica en familias, encontrando que variantes en los genes DRD4, DAT1, DRD5, DBH, SNAP-25 y 5HTT efectivamente se encuentran relacionados con un incremento en la susceptibilidad al desarrollo de este trastorno5.
Uno de los genes candidatos más estudiados es DAT1, que codifica para el receptor del trasportador de dopamina. Su estudio inició debido a la observación de que muchos de los medicamentos usados para el tratamiento del TDAH actúan antagonizando la acción del trasportador de dopamina 1, lo que aumenta la disponibilidad de dicho neurotransmisor en la hendidura sináptica, contribuyendo así a la mejoría de los síntomas6–8. Diferentes estudios han demostrado que polimorfismos del gen DAT1, como la repetición en tándem en número variable (VNTR) de 40 pares de bases (pb) localizado en 3’UTR, pueden ser considerados como un factor de riesgo, debido a que aumentan la expresión del transportador de dopamina9–14. Los genotipos de este VNTR están dados por el número de repeticiones de 40 pb en cada uno de los dos alelos, siendo los de mayor frecuencia a nivel mundial el alelo de 9 repeticiones (alelo 9) y el alelo de 10 repeticiones (alelo 10). El genotipo 10/10, es decir, el genotipo conformado por 2 alelos del 10 repeticiones de 40 pb, se ha encontrado asociado al TDAH. Sin embargo, esta asociación no ha sido replicada en todos los estudios que analizan esta variante genética, por lo que aún no existe un consenso con respecto a la importancia de este polimorfismo en la susceptibilidad al trastorno15–17.
Teniendo en cuenta la discrepancia entre los resultados de estudios que analizan genes de susceptibilidad, se ha propuesto realizar estudios de endofenotipos, que pueden representar alteraciones en la función bioquímica, neurofisiológica o cognitiva y que, indirectamente, son muestra de los procesos fisiopatológicos específicos o subfenotipos de la enfermedad18. Por ejemplo, las alteraciones en la función ejecutiva son frecuentes en pacientes con TDAH y pueden ser consideradas como un endofenotipo del trastorno19. Por este motivo, es posible considerar que su estudio podría llevar a la identificación de variantes genéticas relacionadas específicamente con las alteraciones en las funciones ejecutivas características de este trastorno.
En Colombia, no se han realizado estudios que evalúen la asociación del gen DAT1 con endofenotipos del TDAH. Por consiguiente, el objetivo del presente estudio fue determinar si existe asociación entre el VNTR de 40 pb de la región 3’UTR del gen DAT1 y el fenotipo del TDAH, sus subtipos y endofenotipos (principalmente función ejecutiva) en una muestra de la población colombiana.
Sujetos y métodosSujetosSe estudió a niños entre 6 y 15 años, provenientes de varios colegios públicos y privados de Bogotá, Colombia, a quienes se les aplicó la lista chequeo del Manual Diagnóstico y Estadístico IV (DSM-IV, por sus siglas en inglés), el Sistema de evaluación de la conducta de niños y adolescentes (BASC, por sus siglas en inglés) y la escala Weschler de inteligencia para niños-revisada (WISC-R, por sus siglas en inglés). Los casos fueron seleccionados de acuerdo con los resultados de la lista de chequeo del DSM-IV y la escala BASC. Aquellos niños que cumplieran 6 o más criterios de los 9 para inatención y/o hiperactividad en la lista de chequeo DSM-IV aplicada a padres o maestros y tuvieran un percentil ≥ 85 en el dominio de atención y/o hiperactividad en la escala de atención BASC aplicada a padres o maestros fueron considerados como casos. La prueba WISC-R se aplicó con el fin de excluir de la muestra de estudio a aquellos niños con una puntuación total < 70, concluyente de discapacidad intelectual. Según los resultados, los casos fueron clasificados en subtipos de TDAH mixto, hiperactivo/impulsivo o inatento.
Como controles se tomó a niños en quienes no se sospechaba TDAH y a ellos se les aplicó igualmente la lista de chequeo del DSM-IV y la escala BASC. Para asegurar que los controles no tenían una deficiencia cognitiva se descartaron aquellos que tuvieron puntuación en el WISC-R inferiores a 70 o que tenían un rendimiento escolar bajo (para aquellos a los cuales por cuestiones operativas no se pudo realizar la prueba). Tanto a los casos como a los controles se les realizó una evaluación de las funciones ejecutivas mediante las pruebas de la Evaluación Neuropsicológica Infantil (ENI)20.
Este estudio siguió los principios éticos para la investigación médica que involucra a sujetos humanos emanados de la declaración de Helsinki de la Asociación Médica Mundial (versión Seúl, 2008) y la Resolución N.° 008430 de 1993, por la cual se establecen las normas científicas, técnicas y administrativas para la investigación en salud para Colombia. El protocolo del estudio fue evaluado y aprobado por el Comité de Ética en Investigaciones institucional. Todos los niños participantes (casos y controles) asintieron participar en el estudio y sus padres firmaron un consentimiento informado tanto para la participación como para el uso de sus muestras de ADN.
Extracción de ADNSe obtuvieron 5ml de sangre de vena periférica o muestras de mucosa oral, tanto en los casos como en los controles, dependiendo de la facilidad de la toma de muestra de sangre en los niños. La extracción de ADN fue realizada por métodos convencionales de desalamiento y su calidad fue verificada en geles de agarosa al 1,2% teñidos con bromuro de etidio. La cuantificación del ADN fue realizada utilizando un Nanodrop®. El ADN de casos y controles fue almacenado a –20°C en el banco de ADN de la unidad de genética de la Universidad del Rosario (Bogotá, Colombia) hasta su uso.
GenotipificaciónPara el análisis del polimorfismo de 40 pb del VNTR situado en el extremo 3¿UTR del gen DAT1, se realizó reacción en cadena de la polimerasa (PCR, por sus siglas en inglés), con primers específicos previamente reportados9 que amplificaban un fragmento entre 160 y 640 pb, según el número de repeticiones del VNTR que tuviese cada caso o control, correspondiendo el alelo menor (2 repeticiones) a un peso molecular de 160 pb y el alelo de mayor número de repeticiones (13 repeticiones) a un peso de 640 pb.
La PCR se realizó bajo las siguientes condiciones: Máster Mix (6,25μL Promega®), Primers forward 0,5μL y reverse 0,5μL (10 pm/μl), ADN 150 ng, agua ultrapura 5,25μL, 2μL de dimetil sulfóxido al 10%. El programa de termociclador utilizado consistió en: desnaturalización a 94°C por 5 min, seguido de 30 ciclos de: desnaturalización a 94°C por 45 s, anillamiento a 61°C por 45 s y extensión a 72°C por 45 s, seguidos por una extensión final de 10 min a 72°C (termociclador Agilent®). En cada montaje de PCR se utilizó un control negativo de amplificación para descartar posibles contaminaciones y como controles positivos se usaron muestras previamente tipificadas en la región 3’UTR del gen DAT1. Todos los productos amplificados se analizaron en geles de agarosa al 3%, teñidos con bromuro de etidio al 3%, utilizando un marcador de peso molecular de 50 pb. La genotipificación fue realizada mediante visualización directa del producto amplificado en el gel, por conteo directo de los alelos y por 2 coinvestigadores en evaluaciones independientes. La asignación del genotipo fue realizada por comparación de la banda de amplificación obtenida con el patrón de peso molecular, teniendo en cuenta que el alelo de menor número de repeticiones (2 repeticiones) tiene un peso de 160 pb.
Análisis estadísticoSe utilizó el paquete estadístico SNPStats21 para determinar los estadísticos genético-poblacionales de frecuencias alélicas, frecuencias genotípicas y equilibrio de Hardy-Weinberg. Se calcularon las OR del genotipo vs. fenotipo y subtipos de TDAH. Usando el programa estadístico SPSS20, se realizaron análisis bivariados mediante pruebas de correlación entre el genotipo del VNTR situado en el extremo 3¿UTR del gen DAT1 y los resultados obtenidos en las subpruebas del WISC y de las funciones ejecutivas de la ENI (fluidez, fluidez gráfica, flexibilidad cognitiva, planificación y organización), las pruebas de memoria y el test de la figura humana. Las resultados se evaluaron con una tasa de error del 5% (valor de p < 0,05).
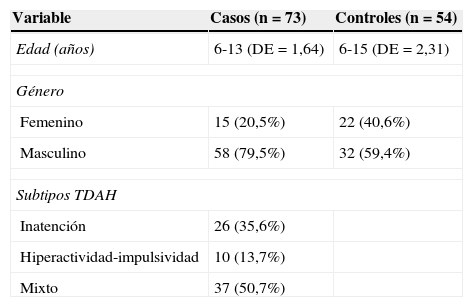
ResultadosSe analizó a 73 niños con diagnóstico de TDAH cuyas edades se encontraban entre los 6 y los 13 años (DE=1,64) y 54 controles sanos, con edades entre los 6 y los 15 años (DE=2,31). Los casos fueron clasificados en 3 diferentes subtipos, de acuerdo con los resultados en la clasificación DSM-IV y el BASC. La mayoría de los casos correspondió al subtipo mixto. La relación entre hombres y mujeres en el grupo de casos fue de ∼ 4:1 (tabla 1).
No fue posible la genotipificación de 4 de las muestras obtenidas (debido a bajas concentraciones del ADN disponible), correspondientes a un control y 3 casos, 2 de ellos del subtipo inatento y uno del subtipo hiperactivo.
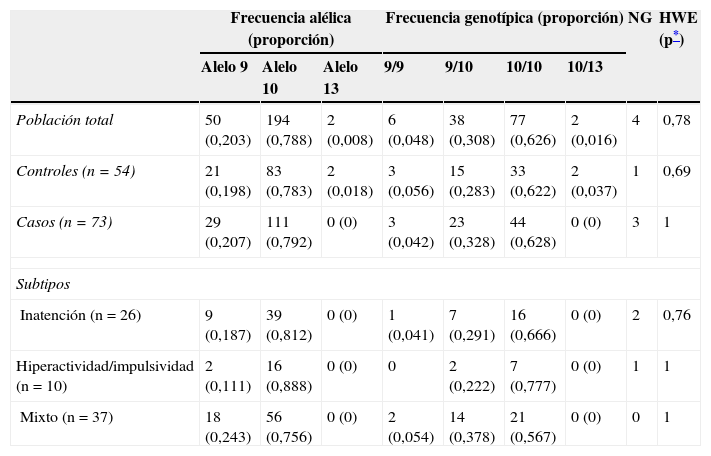
No hubo diferencias significativas en frecuencias alélicas y genotípicas de casos y controles (p=0,26 y p=0,39, respectivamente) (tabla 2). La población se encontró en equilibrio de Hardy Weinberg (p=0,78). El genotipo más frecuente, tanto en los casos como en los controles, fue 10/10 (2 alelos de 10 repeticiones), con frecuencias del 62,8 y el 62,2%, respectivamente.
Valores observados y proporción de las frecuencias alélicas y frecuencias genotípicas de casos y controles
| Frecuencia alélica (proporción) | Frecuencia genotípica (proporción) | NG | HWE (p*) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Alelo 9 | Alelo 10 | Alelo 13 | 9/9 | 9/10 | 10/10 | 10/13 | |||
| Población total | 50 (0,203) | 194 (0,788) | 2 (0,008) | 6 (0,048) | 38 (0,308) | 77 (0,626) | 2 (0,016) | 4 | 0,78 |
| Controles (n = 54) | 21 (0,198) | 83 (0,783) | 2 (0,018) | 3 (0,056) | 15 (0,283) | 33 (0,622) | 2 (0,037) | 1 | 0,69 |
| Casos (n = 73) | 29 (0,207) | 111 (0,792) | 0 (0) | 3 (0,042) | 23 (0,328) | 44 (0,628) | 0 (0) | 3 | 1 |
| Subtipos | |||||||||
| Inatención (n = 26) | 9 (0,187) | 39 (0,812) | 0 (0) | 1 (0,041) | 7 (0,291) | 16 (0,666) | 0 (0) | 2 | 0,76 |
| Hiperactividad/impulsividad (n = 10) | 2 (0,111) | 16 (0,888) | 0 (0) | 0 | 2 (0,222) | 7 (0,777) | 0 (0) | 1 | 1 |
| Mixto (n = 37) | 18 (0,243) | 56 (0,756) | 0 (0) | 2 (0,054) | 14 (0,378) | 21 (0,567) | 0 (0) | 0 | 1 |
HWE: equilibrio de Hardy Weinberg; NG: no genotipificación.
No se obtuvo una asociación entre el genotipo del VNTR del 3¿UTR del gen DAT1 y el fenotipo de TDAH (p=0,85). Tampoco se encontró asociación con los subtipos de inatención, hiperactividad/impulsividad, mixto (p=0,83, p=0,40 y p=0,44, respectivamente).
Al realizar el estudio de correlación entre las subpruebas tanto del estudio del WISC como de las funciones ejecutivas con el genotipo del VNTR del 3¿UTR del gen DAT1, se evidenció solo una correlación estadísticamente significativa entre el genotipo y el índice de velocidad de procesamiento del WISC (p=0,043) (tabla 3). Se determinó que los casos portadores del genotipo 10/10 tienen medias más bajas en este índice, comparado con otros genotipos (media del genotipo 10/10=88; media del genotipo 9/9 y 9/10=95).
Correlaciones de endofenotipos y genotipo del VNTR en 3’UTR del gen DAT1 que presentaron significación estadística
| Fenotipo | Prueba/endofenotipo | R2 ajustado | p* |
|---|---|---|---|
| TDAH | Índice de velocidad de procesamiento (WISC) | 0,046 | 0,043 |
| Hiperactivo-impulsivo | Número de respuestas correctas (flexibilidad cognitiva) | 0,623 | 0,011 |
| Total de errores (flexibilidad cognitiva) | 0,596 | 0,015 | |
| Número de respuestas perseverativas (flexibilidad cognitiva) | 0,753 | 0,001 | |
| Mixto | Índice de velocidad de comprensión (WISC) | 0,096 | 0,037 |
En el subtipo de inatención no se evidenció correlación entre el genotipo y las subpruebas analizadas. Sin embargo, en algunas subpruebas, como la fluidez verbal fonémica total y la fluidez gráfica semántica, se hallaron valores de p cercanos a la significación (p=0,062 y p=0,084, respectivamente). Con respecto al subtipo de hiperactividad-impulsividad, el genotipo de DAT1 se encontró asociado al número de respuestas correctas (p=0,011) y el total de errores (p=0,015) de la subprueba de flexibilidad cognitiva y con el número de respuestas perseverativas (p=0,001). En el subtipo mixto, solo la subprueba índice de comprensión verbal del WISC se asoció significativamente con el genotipo (p=0,037).
Adicionalmente, se realizó un análisis por subgrupos genotípicos con el fin de evaluar la influencia alélica en los endofenotipos. Se agruparon entonces aquellos genotipos que contenían el alelo de 9 repeticiones (es decir, el genotipo 9/9 y el 9/10) y, por otro lado, aquellos genotipos sin el alelo de 9 repeticiones (genotipo 10/10 y 10/13). Mediante este análisis se encontró una relación con la subprueba INVP del WISC (p=0,033), y se demostró una media más alta en este índice (94 vs. 88) en aquellos casos portadores del alelo de 9 repeticiones. No se encontraron otras correlaciones.
DiscusiónEl transportador de dopamina, localizado en la región presináptica en las neuronas dopaminérgicas, regula la recaptación de dopamina dentro de los terminales presinápticos22. El gen DAT1 que codifica para esta proteína ha sido sujeto a numerosos estudios de asociación con TDAH, ya que exhibe varias características para ser un buen gen candidato: ratones «knockout» exhiben un incremento en la actividad motora con residuos de dopamina 100 veces mayores a lo normal en la hendidura sináptica8; estudios de tomografía computarizada de emisión de fotones y positrones, realizados en afectados, revelan mayor densidad de DAT1 a nivel del núcleo estriado23 y es blanco del medicamento metilfenidato, cuyo mecanismo de acción incluye el bloqueo de más del 50% de los transportadores de dopamina a nivel de la neurona presináptica6,24.
Los hallazgos de nuestro estudio indican una relación del genotipo 10/10 en los casos de TDAH con el índice de velocidad de procesamiento, prueba que da una medida acerca de la capacidad de rastrear y discriminar información visual, e informa sobre la atención y la memoria visual a corto plazo. En portadores de este genotipo, se encontraron medias del índice de velocidad de procesamiento más bajas que indican que estos individuos presentan alteraciones en estas habilidades. Igualmente, el análisis por alelos permitió confirmar la correlación de homocigotos para el alelo 10 con el índice de velocidad de procesamiento, lo que indica que este alelo es el que confiere susceptibilidad en la alteración de este endofenotipo, así como lo señalan los hallazgos del estudio de Bellgrove et al., que indican que el alelo de 10 repeticiones puede ser mediador de una alteración neuropsicológica25.
Así mismo, el análisis por subtipos de TDAH permitió evidenciar, en el subtipo de hiperactividad-impulsividad, una relación del genotipo 10/10 con peores resultados en el número de respuestas correctas, el total de errores en la prueba de las cartas y el número de respuestas perseverativas. Estas pruebas en conjunto brindan información sobre la capacidad de anticipación y establecimiento de metas, la autorregulación en las tareas y la capacidad de llevarlas a cabo eficientemente. Además, proporcionan información sobre la capacidad del individuo para adaptarse a los requerimientos del medio ambiente. La alteración en estos endofenotipos estaría en relación con la susceptibilidad de este subtipo a la mala adaptación a los medios que exigen finalización eficaz de tareas y adaptación a medios cambiantes como podrían ser los de un salón de clases.
Por otro lado, el análisis del subgrupo mixto identificó que individuos con genotipo 10/10 en este polimorfismo presentan índices de comprensión verbal más bajos que otros genotipos. Esto indica que el genotipo 10/10 se relaciona con alteraciones en la capacidad verbal de niños con TDAH subtipo mixto. Capilla et al. reportaron que este subtipo presenta alteraciones en la flexibilidad cognitiva a través de evaluaciones con magneto-encefalografía (MEG) mientras realizaban tareas que valoraban esta función26.
De manera similar, otros grupos han encontrado asociación entre el genotipo 10/10 de este polimorfismo y alteración en las pruebas neuropsicológicas. Loo, et al. reportaron una asociación entre el genotipo 10/10 del gen DAT1 y un aumento en el total de errores, además de un incremento en respuestas impulsivas y variabilidad en el tiempo de respuesta27. Bellgrove encontró relación de este genotipo con un pobre rendimiento en las tareas de atención a través de un estudio caso-control, en el que se demostró que los homocigotos para el alelo 10 mostraban mayor variabilidad de respuesta en el test de atención sostenida a las tareas de respuesta e indica que el alelo de 10-repeticiones de DAT1 podría mediar el compromiso neuropsicológico en el TDAH25. En conjunto, estos estudios demuestran la importancia de este polimorfismo en la alteración de las funciones ejecutivas producidas por el TDAH y resaltan la importancia de conocer el trasfondo genético de este desorden, si bien no del trastorno con todos sus criterios, sí de algunas alteraciones neuropsicológicas (endofenotipos) que se podrían ver afectadas en estos pacientes.
En concordancia con nuestros resultados, y como lo hemos expuesto previamente, se ha establecido una asociación de endofenotipos del TDAH con el genotipo 10/10 del VNTR de DAT1. Sin embargo, en algunas poblaciones se han descrito otros alelos del mismo VNTR asociados a TDAH, como el alelo de 7 y 9 repeticiones. Barkley et al. identificaron una relación del genotipo 9/10 con una mayor expresión de los síntomas de TDAH que los portadores del genotipo 10/1028. Adicionalmente, este estudio evidenció que los portadores del genotipo 9/10 tenían peor rendimiento escolar, mayores alteraciones en la relación madre-hijo y mayores alteraciones comportamentales. También Kim et al. encontraron que el genotipo 9/10 era más frecuente en los sujetos con TDAH y las frecuencias alélicas del alelo 9 repeticiones eran significativamente más altas en los sujetos con TDAH29. El autor indica que los sujetos portadores del alelo de 9 repeticiones cometen más errores por omisión en las tareas de vigilancia continua. Por su parte, Oh et al. encontró que los individuos con el genotipo 10/10 cometen menos errores de omisión en el test de variables de atención que los de genotipo 10/* en el primer cuarto del test, lo que indicaría un factor protector para los individuos con el genotipo 10/1030.
Estas observaciones que asocian genotipos diferentes del 10/10 con el TDAH en diferentes poblaciones podrían ser explicadas por varios aspectos: diferencias en el valor de heredabilidad entre las poblaciones; impacto del ambiente psicosocial como factor importante de riesgo; desequilibrio de ligamiento entre el VNTR analizado y otros polimorfismos funcionalmente responsables del fenotipo; influencia de haplotipos más que de polimorfismos únicos en la susceptibilidad al fenotipo y tipo de análisis realizado, ya que en estudios caso-control se puede presentar estratificación de población y en los análisis de test de desequilibrio de transmisión puede influir la homocigocidad de los padres y la fenotipificación de pacientes. Se ha establecido también que, dada la cantidad de estudios que no demuestran asociación de DAT1 con TDAH, a pesar de ser biológica y fisiológicamente un candidato óptimo, este gen podría corresponder más a un modulador de fenotipo de acción dependiente de la interacción con otros polimorfismos de vías dopaminérgicas como DRD415,17.
Los hallazgos reportados en este estudio demuestran la importancia de conocer el genotipo del VNTR localizado en la región 3’UTR del gen DAT1 en poblaciones colombianas, ya que los individuos portadores del genotipo 10/10 se relacionan con un peor desempeño en pruebas neuropsicológicas.
El presente estudio tiene algunas limitaciones referentes al reducido tamaño de muestra, lo que influye en la generalización y la validez externa de los resultados, y a que la población de estudio se ubicó en algunos colegios de ciudad de Bogotá, por lo que estos resultados deben ser tomados cuidadosamente en otras poblaciones. En estudios futuros se recomienda ampliar el tamaño de la muestra, tanto local como regionalmente, especialmente en aquellas con datos de una mayor prevalencia del trastorno, lo que permitiría la identificación de alelos de baja frecuencia, que son difíciles de detectar en bajos tamaños poblacionales2; evaluar otras variantes genéticas del gen DAT1 y otros genes implicados que podrían estar enmascarando relaciones reportadas en estudios previos y, finalmente, realizar estudios funcionales que permitan dilucidar con mayor claridad el papel biológico que cumple este polimorfismo en la alteración de las pruebas neuropsicológicas mencionadas.
FinanciaciónEste proyecto fue financiado por el Fondo de Investigaciones de la Universidad del Rosario (FIUR).
Conflicto de interesesLos autores declaran no tener ningún conflicto de interés.
Agradecemos a las instituciones educativas participantes de este proyecto y a la red de colegios de Colsubsidio, por su disposición e interés en el buen desarrollo del mismo.
Este trabajo no ha sido presentado en la reunión anual de la SEN ni en otras reuniones o congresos.

