





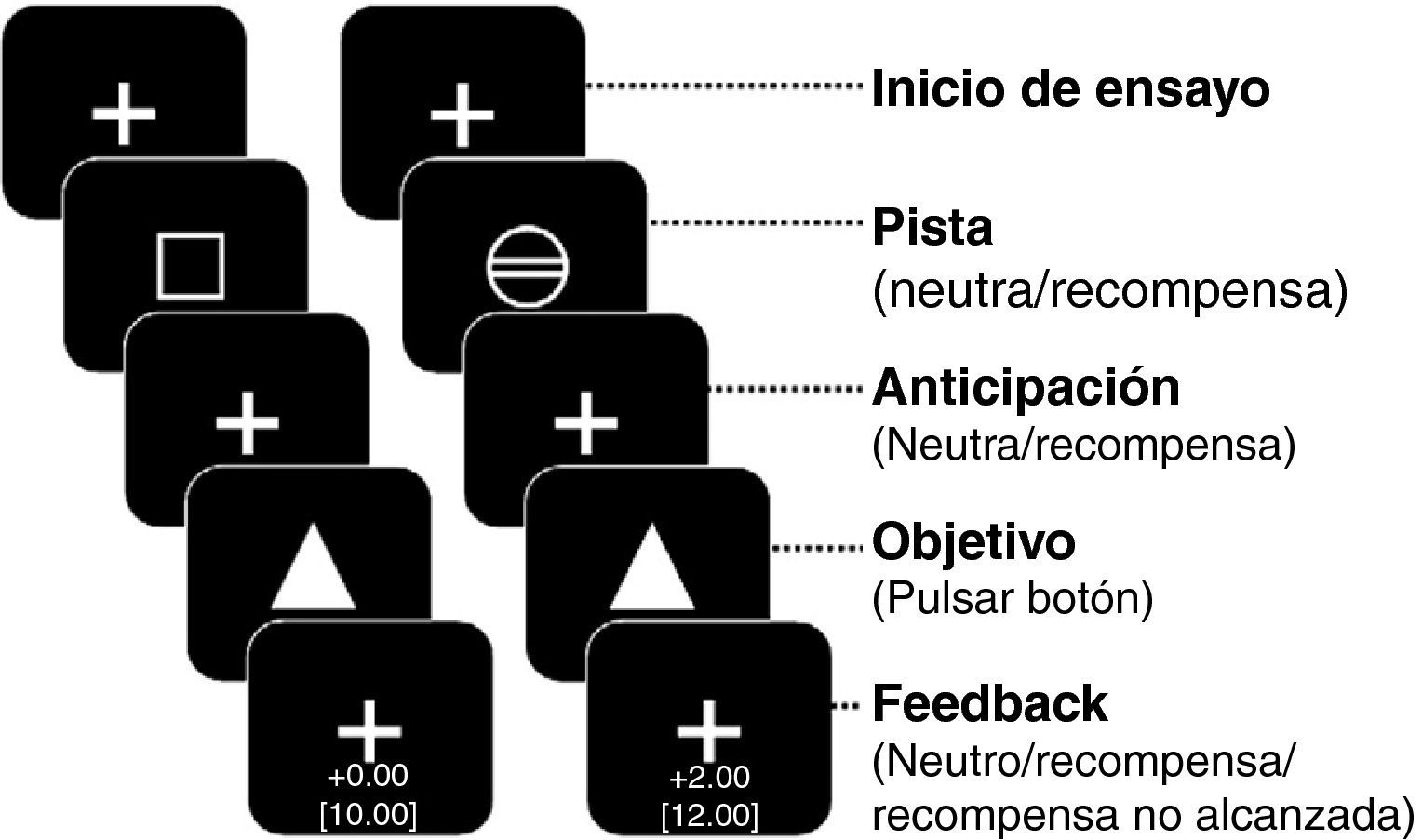
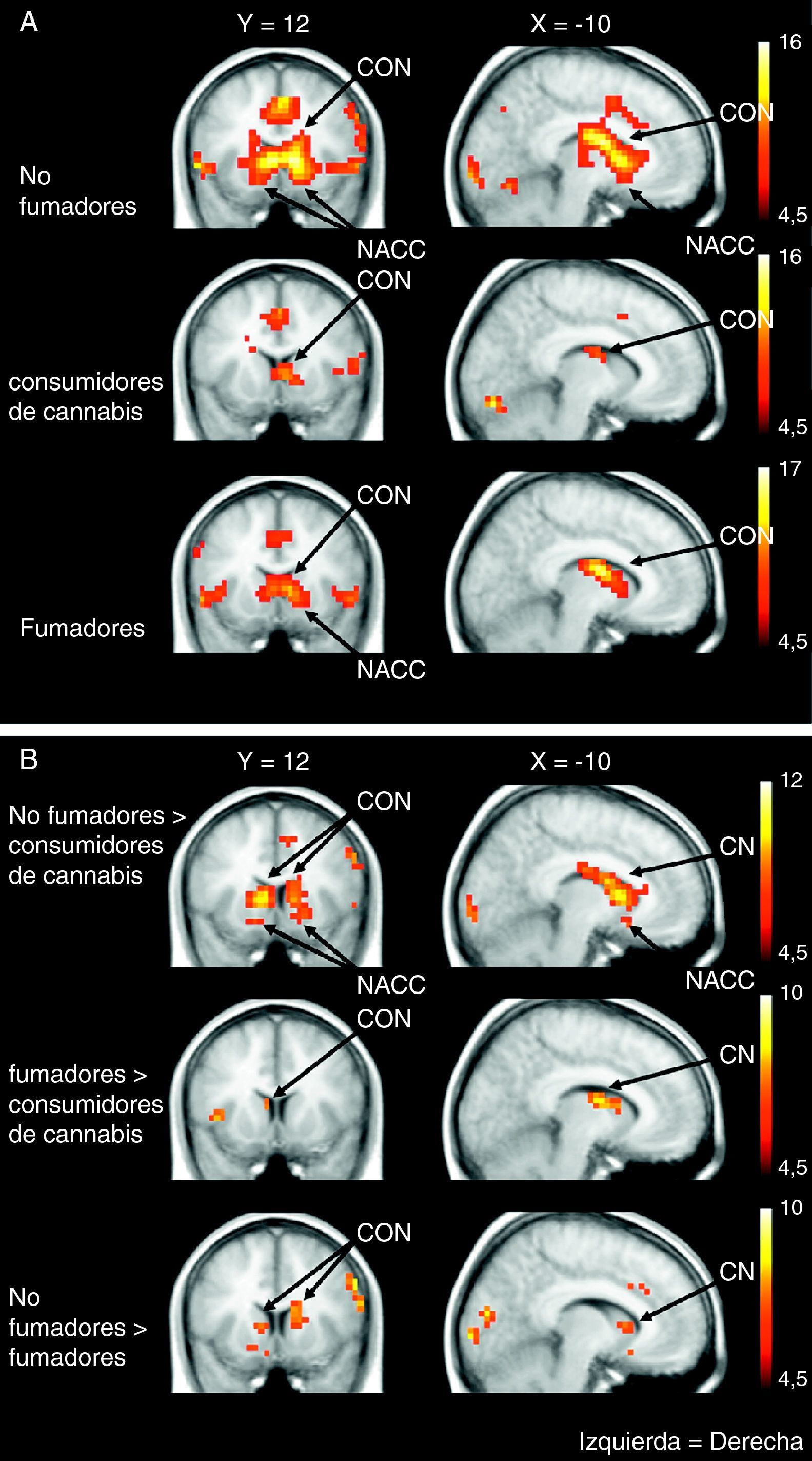
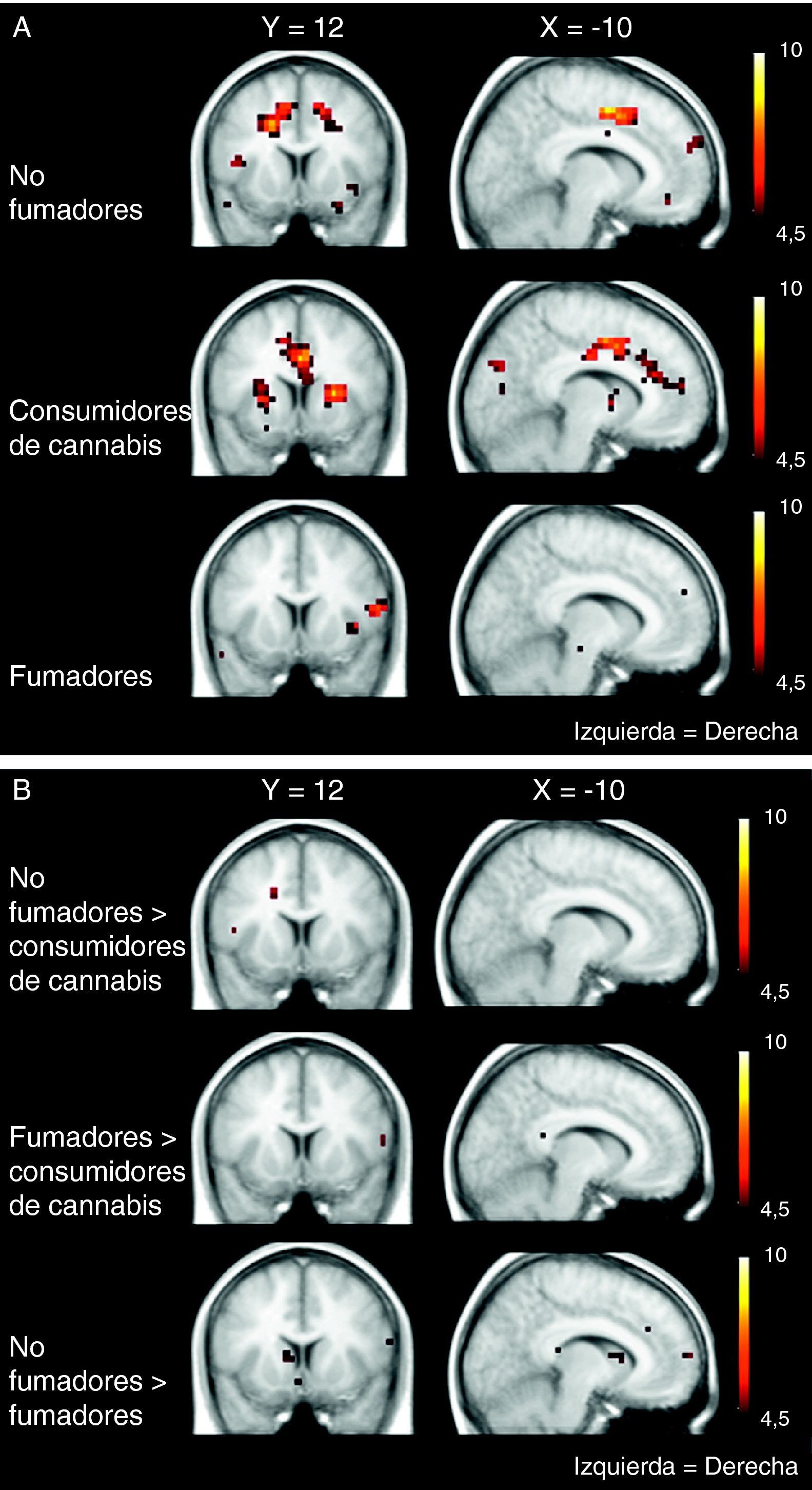
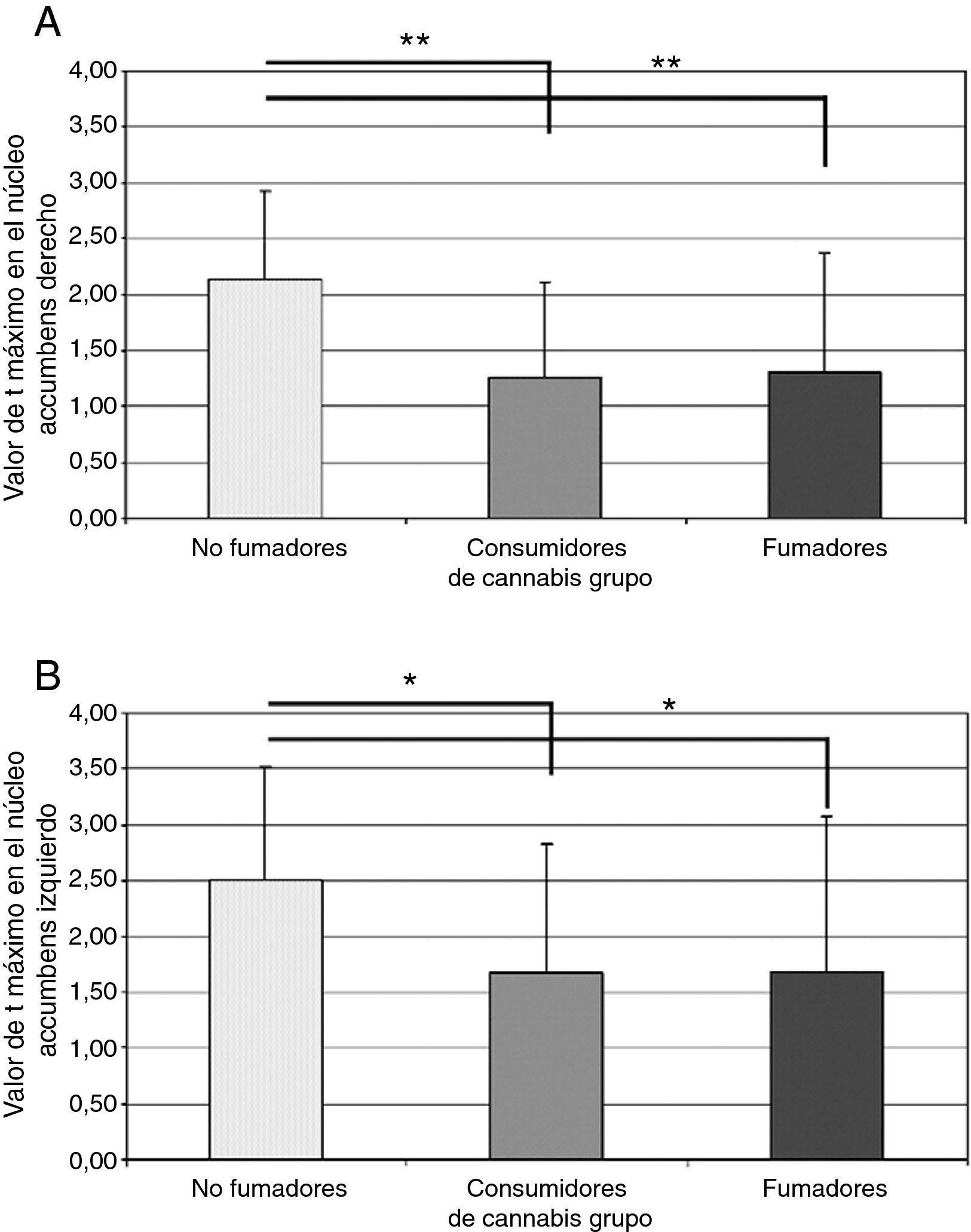
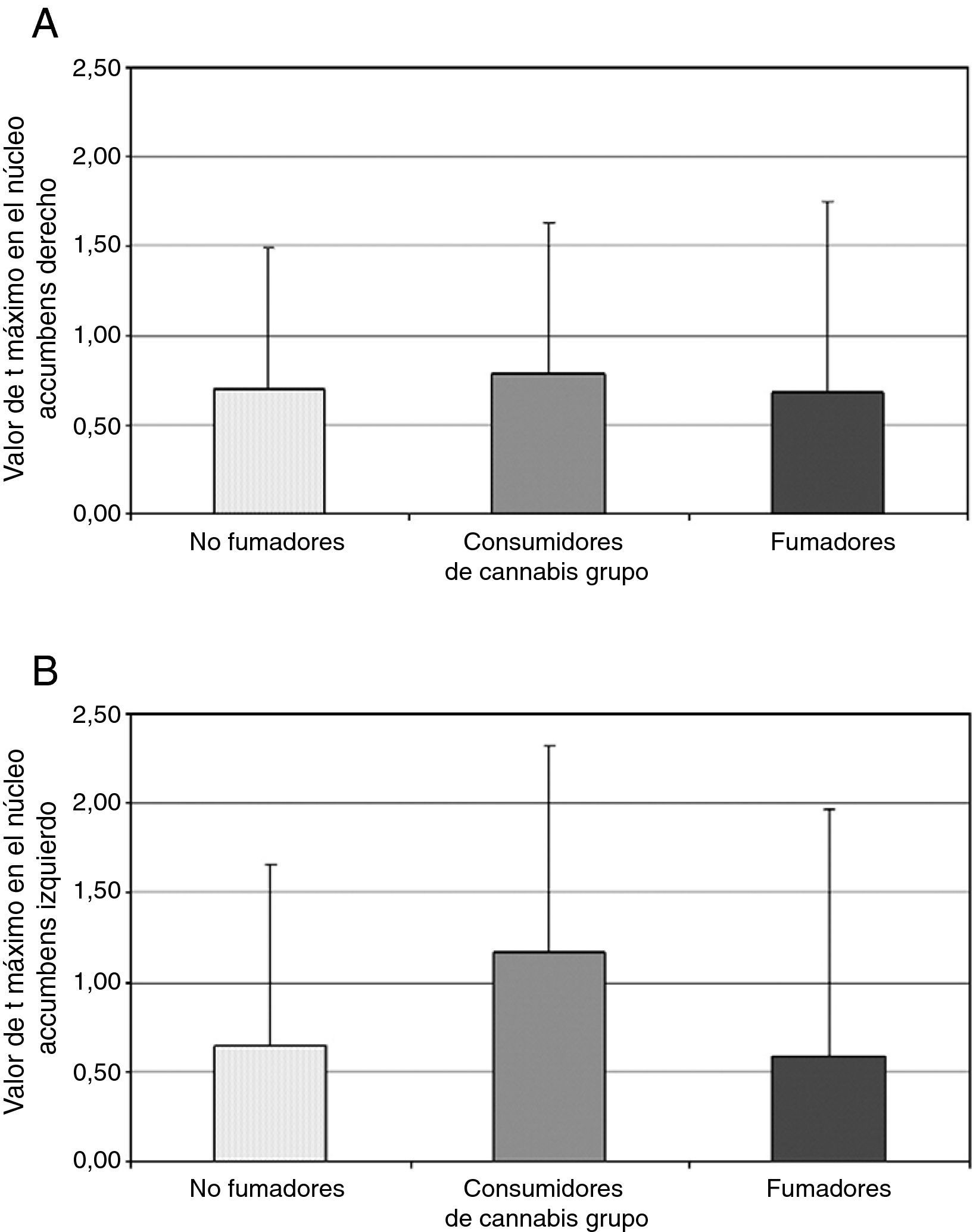
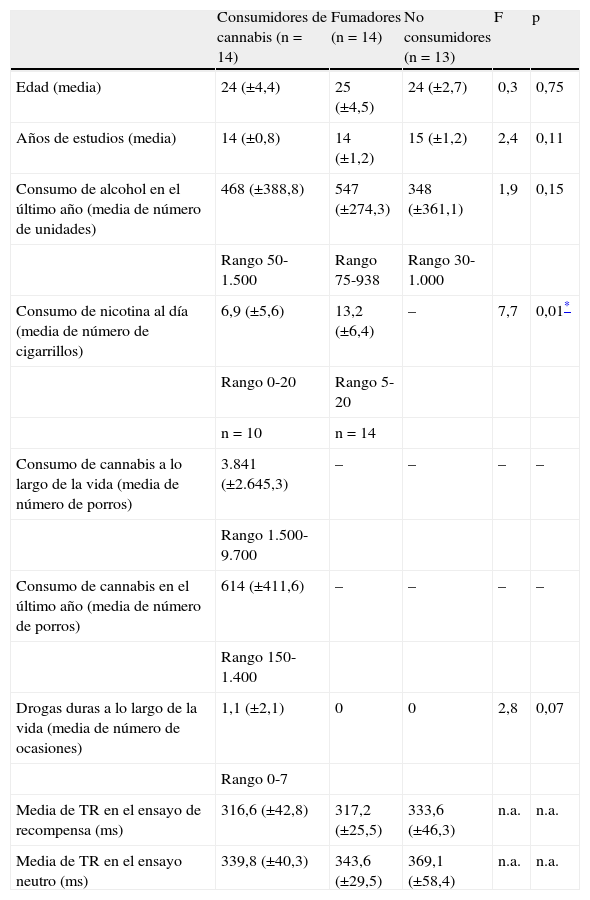
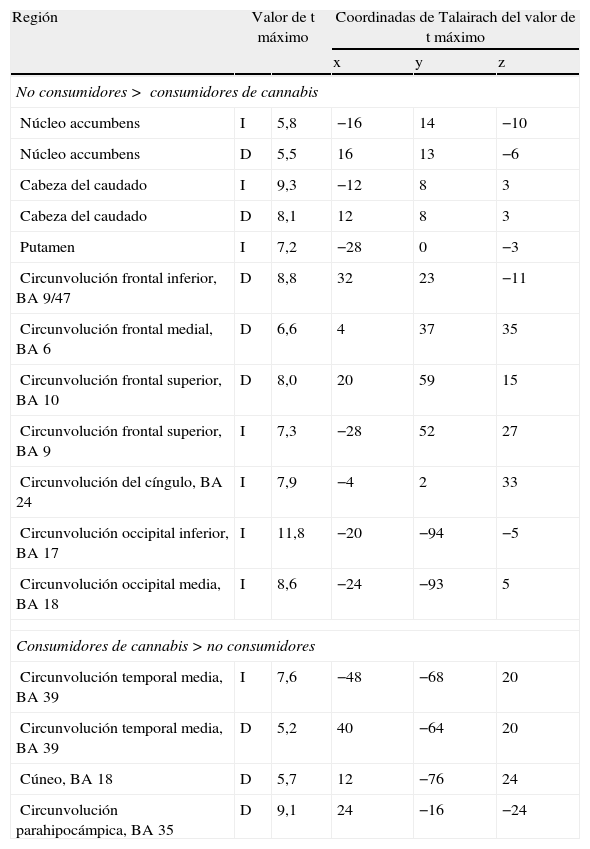
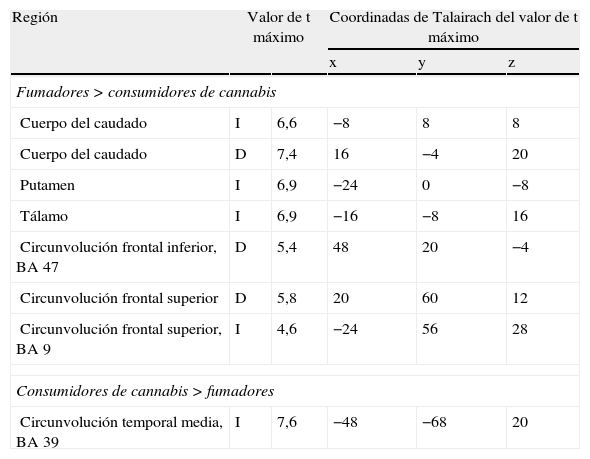
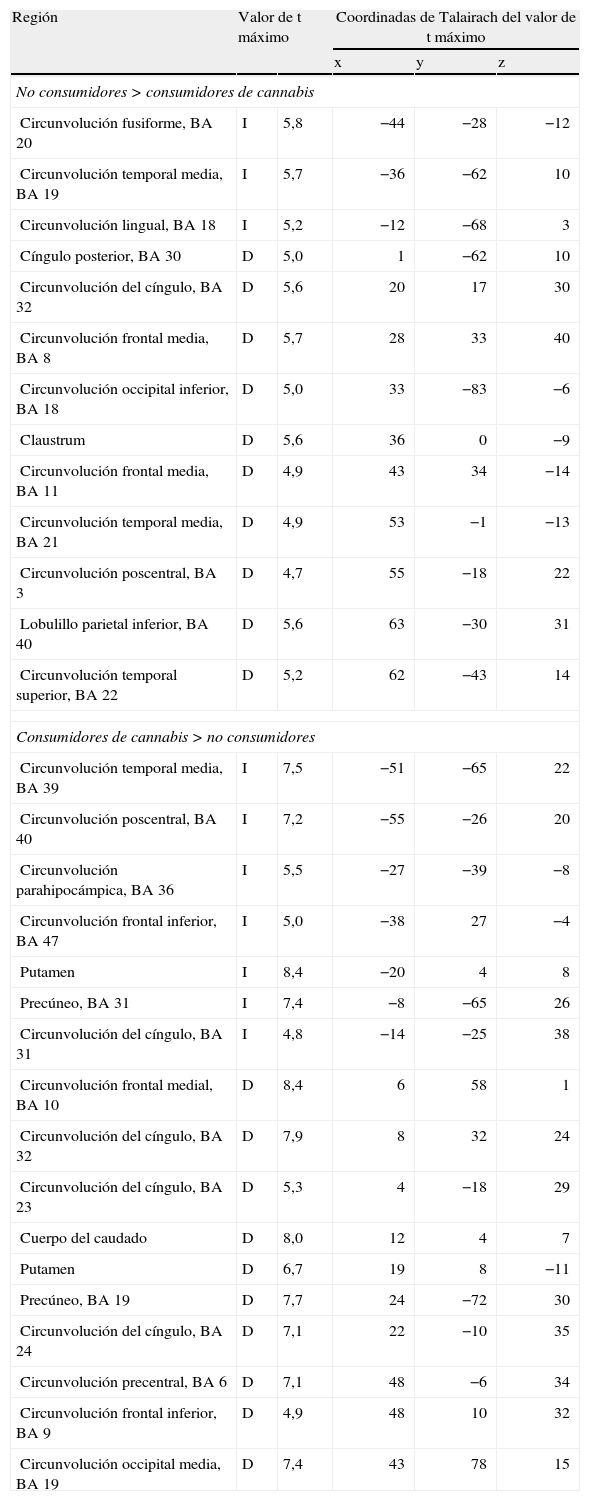
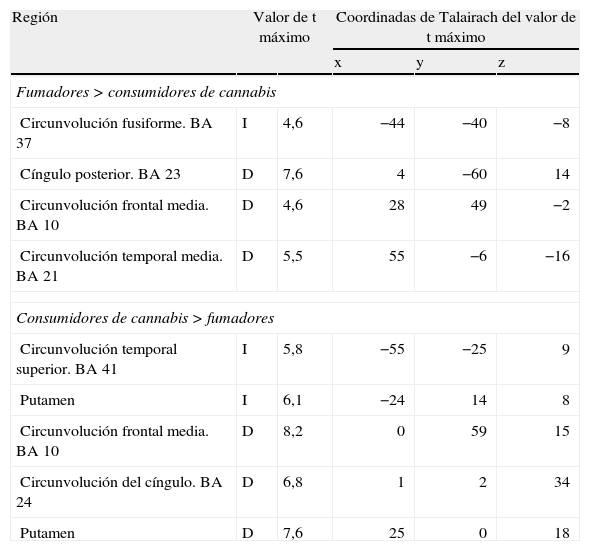
El cannabis es una de las drogas de abuso más frecuentes. Afecta al sistema de recompensa cerebral en los animales, y tiene un potencial de recompensa y adicción demostrado en el ser humano. Hemos utilizado la RM funcional para medir la actividad cerebral durante la anticipación de la recompensa en una tarea de recompensa monetaria. Se comparó a consumidores crónicos de cannabis con individuos de control sanos. Se utilizó otro grupo control adicional formado por consumidores de nicotina. Los consumidores de cannabis mostraron una actividad cerebral atenuada durante la anticipación de la recompensa en el núcleo accumbens, en comparación con los controles no fumadores, pero no en comparación con los controles fumadores. Los consumidores de cannabis mostraron una reducción de la actividad de anticipación de recompensa en el núcleo caudado, en comparación con los controles tanto fumadores como no fumadores. Estos datos sugieren que la nicotina puede ser responsable de una atenuación de la actividad de anticipación de recompensa en el núcleo accumbens, pero que las diferencias que se producen en el caudado se asocian al consumo de cannabis. Nuestros resultados implican que el consumo crónico de cannabis, así como el de nicotina, puede causar una alteración de la respuesta cerebral a los estímulos de recompensa.
Cannabis is one of the most used drugs of abuse. It affects the brain reward system in animals, and has proven rewarding and addictive potential in humans. We used functional MRI to measure brain activity during reward anticipation in a monetary reward task. Long-term cannabis users were compared to healthy controls. An additional control group consisting of nicotine users was included. Cannabis users showed attenuated brain activity during reward anticipation in the nucleus accumbens compared to non-smoking controls, but not compared to smoking controls. Cannabis users showed decreased reward anticipation activity in the caudate nucleus, compared to both non-smoking and smoking controls. These data suggest that nicotine may be responsible for attenuated reward anticipation activity in the accumbens, but that differences in the caudate are associated with the use of cannabis. Our findings imply that chronic cannabis use as well as nicotine, may cause an altered brain response to rewarding stimuli.
Artículo
Comprando el artículo el PDF del mismo podrá ser descargado
Precio 19,34 €
Comprar ahora