





Agriculture is one of the bases of the Argentine economy. Glyphosate is undoubtedly one of the most important herbicides used. The increasing consumption and the efficiency of glyphosate-based herbicides have encouraged several studies on their persistence in soils, their effects on soil microbiota and their degradation processes. Fungi have been reported as being the main herbicide-degrading microorganisms as well as the most tolerant to environmental stress conditions. This study evaluated the growth performance of Aspergillus section Flavi and Aspergillus niger aggregate strains on Czapek Dox media supplied with a commercial glyphosate formulation as sole source of carbon (CZC), phosphorus (CZP) or nitrogen (CZN). Six Aspergillus spp. strains were evaluated. Each medium was stab-inoculated with fungal spores from 7-day old cultures. Two measures of colony radii were taken daily. All of the Aspergillus section Flavi strains showed a significant increase (from 24 to 44%) in growth rate on the CZN medium, as compared to controls. The A. niger aggregate strains exhibited the same behavioral pattern under all the conditions tested, except on the CZN medium. Velutinous or slightly floccose colonies with abundant sporulation were observed on CZP. Moreover, the colonies produced sparse sporulation on CZC or CZN media, being their appearances completely different from those on the CZP medium. This study establishes that A. section Flavi and A. niger aggregate strains can grow in vitro in the presence of glyphosate, especially when it is used as a sole source of phosphorus or nitrogen.
La agricultura es una de las bases de la economía argentina. El glifosato es, indudablemente, uno de los herbicidas más utilizados. El incremento en el consumo y la eficiencia de estos herbicidas han estimulado investigaciones sobre su persistencia en el suelo, sus efectos sobre la microbiota edáfica y su proceso de degradación. Los hongos son conocidos por su importancia como degradadores de herbicidas y por su tolerancia a condiciones de estrés ambiental. El objetivo del presente estudio fue evaluar el crecimiento de cepas de Aspergillus sección Flavi y del agregado Aspergillus niger en el medio Czapeck Dox suplementado con una solución comercial de glifosato como única fuente de carbono (CZC), de fósforo (CZP) o de nitrógeno (CZN). Seis cepas de Aspergillus fueron evaluadas. Cada medio fue inoculado con esporas fúngicas provenientes de cultivos de 7 días. Diariamente se tomaron 2 medidas del radio de las colonias. Todas las cepas de Aspergillus sección Flavi mostraron un incremento significativo (del 24 al 44%) en la velocidad de crecimiento en el medio CZN comparado con los controles. Las cepas del agregado A. niger mostraron el mismo comportamiento bajo todas las condiciones ensayadas, excepto en el medio CZN. Se observaron colonias velutinosas o ligeramente flocosas con abundante esporulación en los medios CZP. En los medios CZC o CZN, las colonias produjeron una esporulación escasa. Este estudio establece que cepas de Aspergillus sección Flavi y del agregado A. niger pueden desarrollar in vitro en presencia de glifosato, especialmente cuando es usado como única fuente de fósforo o nitrógeno.
Agriculture is one of the bases of the Argentine economy. Some of the country's main exports are soybean (Glycine max L.) and maize (Zea mays L). During the last year, the estimated total areas planted with soybean and maize were of 20350000 and 3300000ha, respectively8. Since the 1990s, a significant agricultural transformation has been taking place in the country. Such transformation has been motorized by the adoption of transgenic crops, no-tillage systems and the use of chemical products to protect plants from pests and diseases28.
This new intensive agricultural system duplicated Argentinean production during the past decade. In this context, glyphosate (N-phosphonomethylglycine) is undoubtedly one of the most important herbicides used24. While the normal dose for glyphosate is 4–5.5l/ha, equivalent to 2–2.75kg/ha of glyphosate27, repeated applications are commonly used in no-tillage systems25. Significant amounts of glyphosate (among other pesticides) are applied annually. However, it is widely believed that only a small fraction of these products effectively reaches the target organisms, and that the surpluses are being introduced inadvertently into the soil contaminating non-target organisms while moving into the atmosphere and the water16. The widespread incorporation of herbicides into soils every year has a significant impact on their quality, and on both decomposition and nutrient cycling. This fact has generated great concern for the potential threat that herbicides represent for animal and human health14,36. The increasing consumption and the high popularity and efficiency of glyphosate-based herbicides have encouraged several studies on their persistence in soils, their effects on soil microbiota and their biodegradation processes. Fungal strains have been reported as being the main herbicide-degrading microorganisms as well as the most tolerant to environmental stress conditions. They can produce high amounts of extracellular enzymes during hyphal colonization of soil, resulting in enhanced xenobiotic biodegradation rates26.
Glyphosate degradation in the soil environment is a co-metabolic process and the decomposition rate should depend on the general activity of microbial populations, soil type and environmental conditions37,1. Two major pathways of degradation have been described. One pathway causes the formation of sarcosine and inorganic phosphate via a C-P lyase, while the other type of degradation occurs by a glyphosate oxidoreductase (GOX), which breaks off the glyphosate C-N bond to produce aminomethylphosphonic acid (AMPA) and glyoxylate15. The research on the microbial utilization of glyphosate has tended to focus on the bacterial degradation of this compound. Only a limited number of studies have reported that fungal species are capable of degrading organophosphate compounds such as glyphosate and developing in culture media containing this herbicide mainly as a source of carbon and phosphorus. Among the studied fungal species are Aspergillus flavus, Aspergillus sydowii, Aspergillus niger, Trichoderma viride, Penicillium waksmanii, Penicillium lilacinum, Mucor spp., Fusarium spp. and Saccharomyces rouxii10,13,19,20,21,22,31,38,39,42. A. flavus and A. niger strains are saprophytic soil borne fungi that commonly infect and contaminate vegetables, cereals and oilseed grains at pre- and post-harvest stages30. In a previous study, Carranza et al.11 evaluated the culturable mycobiota of agricultural soils and showed that A. flavus and A. niger aggregate strains are the most prevalent species. A. flavus strains were isolated in percentages ranging from 50% to 87.5% in all soils followed by A. niger aggregate strains isolated in percentages of 50% from corn and soybean soils and 33% from corn-soybean rotation soils. In addition, the effect of different glyphosate concentrations on the growth rate of toxigenic and nontoxigenic Aspergillus section Flavi strains isolated from these soils was evaluated in vitro under different water availability conditions7,11. In view of the mentioned results and considering that glyphosate is not applied to the fields in its pure form, investigating the impact of one of the most used glyphosate-based commercial formulations on fungi may be a more relevant approach. In addition, there is no research on the utilization of this pesticide as nutrient by A. flavus and A. niger aggregate strains. This study was undertaken to evaluate the effects of glyphosate as sole source of carbon, phosphorus or nitrogen on growth parameters and macroscopic colony characteristics of these fungi in vitro.
Materials and methodsFungal strainsSix Aspergillus spp. strains were evaluated. Four Aspergillus section Flavi strains: A. oryzae (AM 1, AM 2, GM 3), A. flavus (GM 4) and two A. niger aggregate strains (AN 251 and AN 384). These strains were previously isolated from fields destined to maize and soybean production, which are located in the south of the province of Córdoba, Argentina. These fields have been exposed to successive applications of pesticides during the last decade11. The strains were identified by traditional taxonomy according to Pitt and Hocking30 and Samson et al.34 for Aspergillus section Flavi and Samson et al.33 for Aspergillus section Nigri strains. The ability of Aspergillus section Flavi strains to produce aflatoxins (AFs) and cyclopiazonic acid (CPA) as well as the ochratoxigenic capacity of A. niger aggregate strains had been previously reported6,11. Only the nontoxigenic strains were selected for this study. The strains belong to our culture collection of the Department of Microbiology and Immunology, at the National University of Río Cuarto, Córdoba, Argentina, and are maintained in 15% glycerol (Sigma–Aldrich, St. Louis, MO, USA) at −80°C.
Culture mediaThe Czapek Dox medium (sucrose 30g, NaNO3 3g, K2HPO4 1g, MgSO4·7H2O 0.25g, KCl 0.5g, FeSO4·7H2O 0.01g, agar 15g, distilled water 1l) was used as control. In addition, sucrose was replaced by glyphosate at a final concentration of 10mM as sole carbon source. When glyphosate was used as sole source of phosphorus or nitrogen, K2HPO4 and NaNO3 were replaced by glyphosate at final concentrations of 1.0 and 1.5mM, respectively22. The control Czapek Dox agar is referred to as CZ. The growth medium in which glyphosate replaces the organic source of carbon is referred to as CZC. Furthermore, when the mineral phosphate or nitrogen sources are replaced by this pesticide the media are referred to as CZP or CZN, respectively. The water agar (WA) medium was also used as control.
GlyphosateThe glyphosate (N-phosphonomethylglycine) used in this study was obtained from a commercial formulation (Round-up1®; Monsanto, Buenos Aires, Argentina), corresponding to a 3.5M solution of the active ingredient of glyphosate (588g/l). A stock solution of glyphosate (0.5M) was prepared by dissolving 14.4ml of the commercial formulation in 100ml of sterile distilled water (v/v). This stock solution was sterilized using 0.2μm filters (Whatman) and kept at 4°C. The appropriate concentration of commercial herbicide solution was applied to the sterilized culture media (tempered at 45–50°C) to obtain the required final concentrations of glyphosate (1.0, 1.5 and 10mM). 20ml of media were poured into 90-mm sterile Petri dishes. These concentrations were selected based on the percentages that the carbon, nitrogen or phosphorus sources should represent in the fungal media. They are below the recommended glyphosate field application rates used (2–2.75kg/ha)27.
Inoculation, incubation and growth parametersEach plate was stab-inoculated in the center, using a sterile loop, with fungal spores harvested from 7-day old cultures grown on malt extract agar (MEA) and suspended in 0.2% of soft agar. Four replicate plates were used per medium. The plates were incubated at 25°C for 28 days.
Two measures of colony radii, at right angles to each other, were taken daily from each replicate. The average radius of each colony was plotted against time, and a linear regression was applied in order to obtain the growth rate as the slope of the line to the X-axis. The lag phase (h) before growth was also determined5.
Growth rate and lag phase analyses were performed on four Aspergillus section Flavi and two A. niger aggregate strains, using three culture media supplied with different glyphosate concentrations and the respective controls. Each analysis was carried out in quadruplicate and all the experiments were repeated twice. Additional qualitative observations regarding colony morphology in each culture medium were recorded throughout the experiments.
Statistical analysesThe results obtained in the experiments of replacement effect of the carbon, phosphorus or nitrogen source by glyphosate on Aspergillus section Flavi and A. niger aggregate growth were analyzed by analysis of variance (ANOVA). All data were log10 (x+1) transformed to obtain homogeneity of variance. The means were compared using the Fisher's protected LSD test to evaluate the growth rate and lag phase before growth in the presence of glyphosate as source of carbon, phosphorus or nitrogen in the culture medium. The analyses were conducted using PROC GLM in SAS (SAS Institute, Cary, NC).
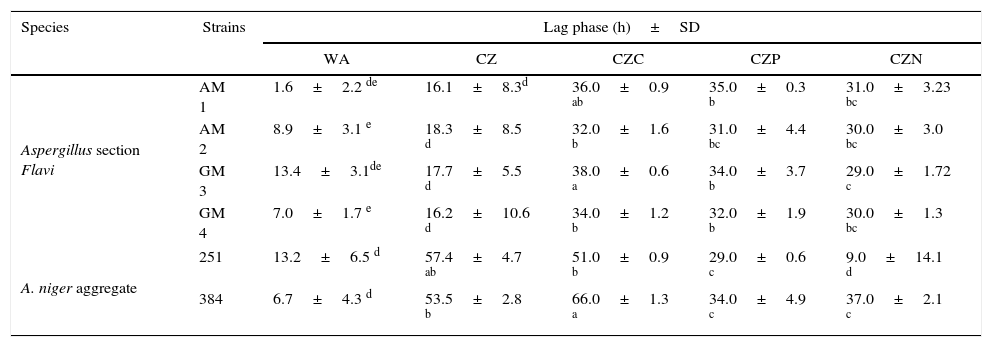
ResultsEffect of the replacement of the carbon, phosphorus or nitrogen source by a commercial glyphosate formulation on lag phasesTable 1 shows the lag phases of four Aspergillus section Flavi and two A. niger aggregate strains in control media and in the presence of different concentrations of glyphosate as the sole source of carbon, phosphorus or nitrogen. In general, all the Aspergillus strains showed the same behavioral pattern in the different media tested. The most noticeable decreases in the lag phases from the Aspergillus section Flavi strains were observed in the WA and control CZ media. Among the supplied media, the shorter lag phase was obtained when glyphosate was included as nitrogen source. In addition, significant differences among the media where glyphosate replaced the carbon, phosphorus or nitrogen source were only found with strain GM 3 (p<0.001). This strain also showed the greatest differences in this parameter between the CZC and CZN media (38 and 29h, respectively).
Lag phases of Aspergillus section Flavi and A. niger aggregate strains in water agar and Czapek Dox agar media supplied with glyphosate
| Species | Strains | Lag phase (h)±SD | ||||
|---|---|---|---|---|---|---|
| WA | CZ | CZC | CZP | CZN | ||
| Aspergillus section Flavi | AM 1 | 1.6±2.2 de | 16.1±8.3d | 36.0±0.9 ab | 35.0±0.3 b | 31.0±3.23 bc |
| AM 2 | 8.9±3.1 e | 18.3±8.5 d | 32.0±1.6 b | 31.0±4.4 bc | 30.0±3.0 bc | |
| GM 3 | 13.4±3.1de | 17.7±5.5 d | 38.0±0.6 a | 34.0±3.7 b | 29.0±1.72 c | |
| GM 4 | 7.0±1.7 e | 16.2±10.6 d | 34.0±1.2 b | 32.0±1.9 b | 30.0±1.3 bc | |
| A. niger aggregate | 251 | 13.2±6.5 d | 57.4±4.7 ab | 51.0±0.9 b | 29.0±0.6 c | 9.0±14.1 d |
| 384 | 6.7±4.3 d | 53.5±2.8 b | 66.0±1.3 a | 34.0±4.9 c | 37.0±2.1 c | |
WA, water agar; CZ, Czapek Dox medium; CZC, Czapek Dox medium without sucrose and supplied with glyphosate at 10mm; CZP, Czapek Dox medium without K2HPO4 and supplied with glyphosate at 1.0mm; CZN, Czapek Dox medium without NaNO3 and supplied with glyphosate at 1.5mm.
Values represent the mean of four replicates. SD: standard deviation. Means from each species with the same letter (a to e) are not significantly different according to the LSD test (p<0.001).
Regarding the A. niger aggregate strains, the lag phase values varied depending on the tested strain. The lowest values were detected in CZN and WA with strains AN 251 and AN 384, respectively. Moreover, the longest lag phases were observed in control CZ (57.4 and 53.5h) and CZC (51.0 and 66.0h) with strains AN 251 and AN 384, respectively (p<0.001).
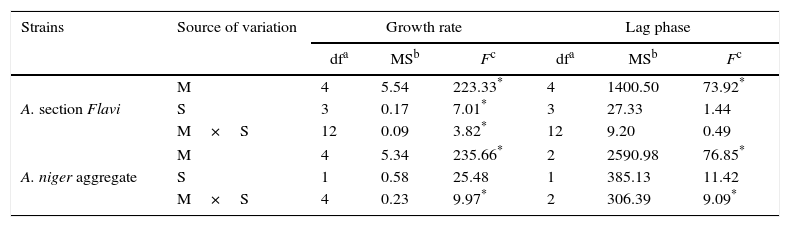
The analysis of variance of the effect of single (medium and strains) and two-way interaction showed that the culture media were statistically significant only in relation to the lag phase for the Aspergillus section Flavi strains. By comparison, the culture media and their interaction were statistically significant for the A. niger aggregate strains (p<0.001) (Table 2).
Variance analysis of effect of medium (M), strains (S) and their interactions on lag phase and growth rate of Aspergillus section Flavi and A. niger aggregate strains in Czapek Dox agar medium supplied with glyphosate
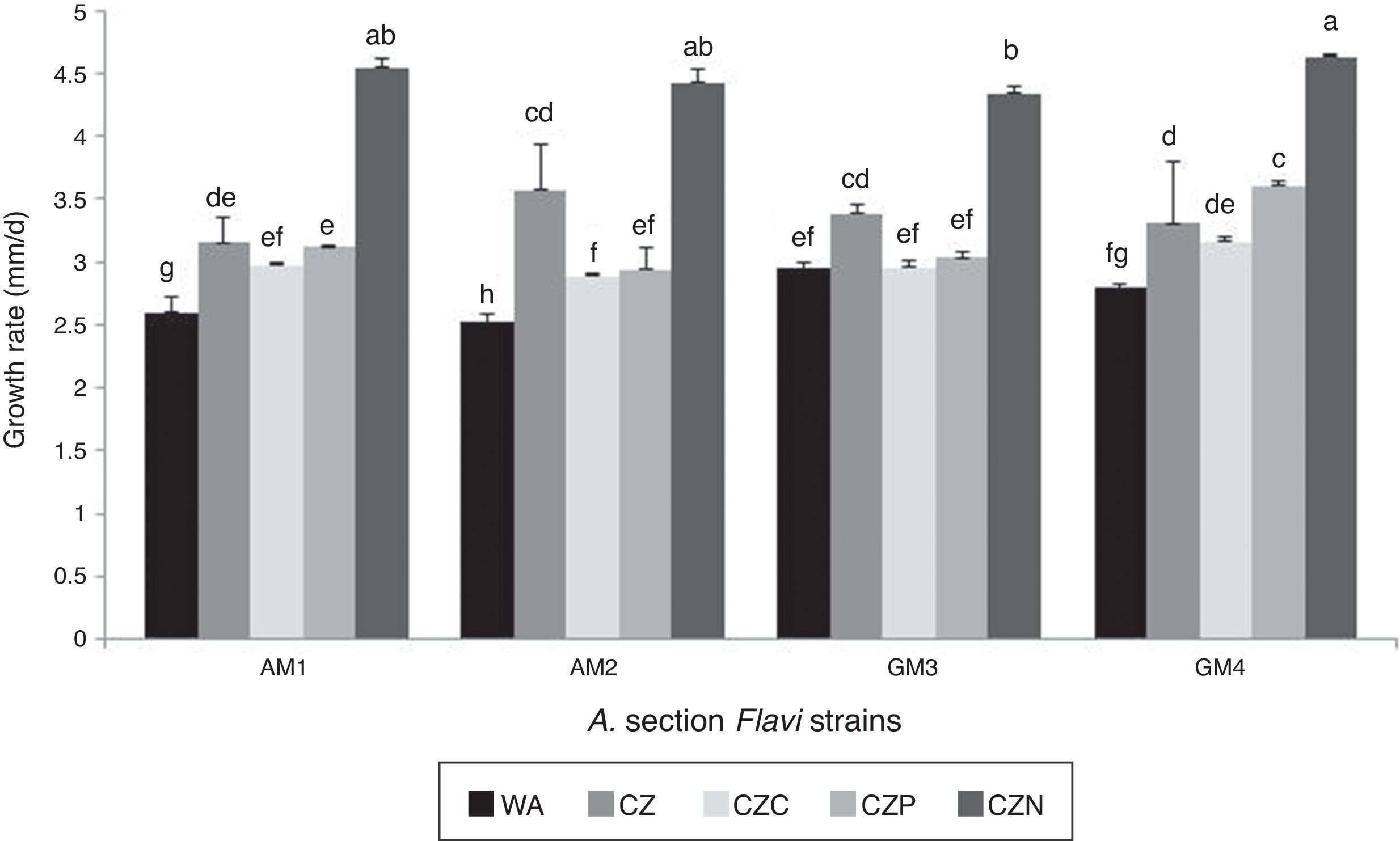
Regarding the growth rate, all Aspergillus section Flavi strains showed a similar behavior. In the medium supplied with glyphosate as nitrogen source, a significant increase (24 to 44%) in growth rate was observed as compared to the control (CZ medium) (p<0.001). In general, no significant differences were detected in this parameter when comparing the media supplied with glyphosate as carbon or phosphorus source. Balanced against each other, significant differences in growth rate values among these media were found in strain GM 4 (p<0.001). The lowest growth rate was obtained in the WA medium (Fig. 1).
.")
Growth rate of Aspergillus section Flavi strains in the presence of a commercial glyphosate formulation as sole source of carbon, phosphorus or nitrogen. Mean values are based on quadruplicated data. Means with a letter in common are not significantly different in accordance with the LSD test (p<0.05).
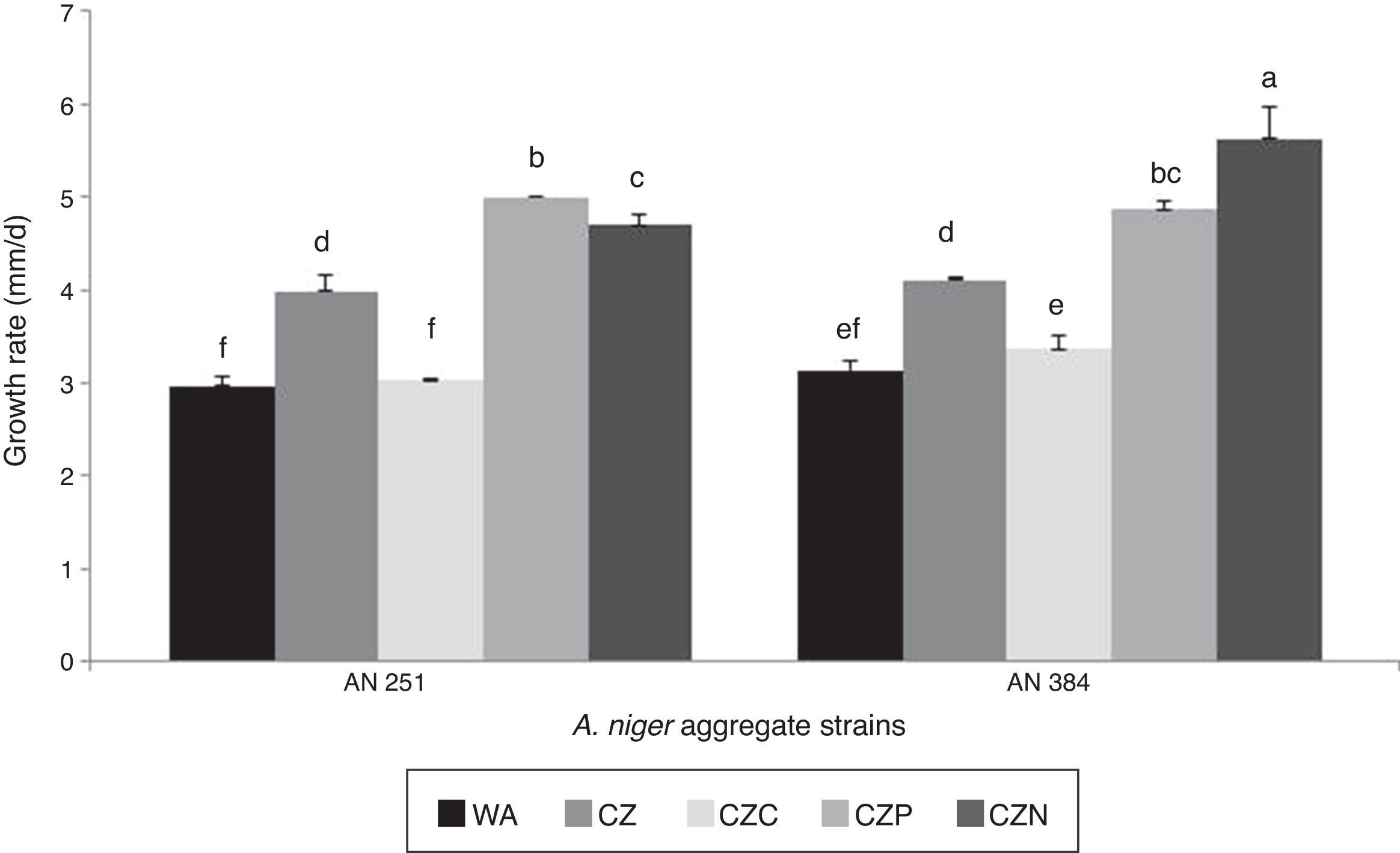
Figure 2 shows the growth rate of two A. niger aggregate strains in control media and in the presence of different concentrations of glyphosate as the sole source of carbon, phosphorus or nitrogen. The two strains showed the same behavioral pattern in all the assayed conditions, except in the media supplied with glyphosate as nitrogen source. In contrast, the two strains showed the lowest values in growth rate on WA and CZC media. Significant differences in growth rates were observed among media amended with glyphosate, as compared to the control (CZ) (p<0.001). In strain AN 251, the highest increase in growth rate (25.5%) compared to the control was found on the CZP medium. Conversely, increases of 19 and 37% in this parameter were observed with strain AN 384 when the pesticide was used to replace the phosphorus and nitrogen sources in the media, respectively.
.")
Growth rate of Aspergillus niger aggregate strains in presence of a commercial glyphosate formulation as sole source of carbon, phosphorus or nitrogen. Mean values are based on quadruplicated data. Means with a letter in common are not significantly different in accordance with the LSD test (p<0.05).
The analysis of variance of the growth rates showed different results depending on the tested species. The effects of each single variable (medium and strains) and the two-way interaction were statistically significant for the Aspergillus section Flavi strains. The culture media and their interaction were statistically significant for the A. niger aggregate strains (p<0.001) (Table 2).
Macroscopic characteristics of colonies in culture media with a commercial glyphosate formulation as sole source of carbon, phosphorus or nitrogenThe macroscopic changes of the morphology of colonies were examined in different media. Changes were similar in all the evaluated strains (data no shown). Colonies from Aspergillus section Flavi strains were plane with abundant white mycelium on the margins on the CZC medium. In addition, abundant sporulation was only observed in the center of the colonies. Their coloration was olive green, similar to that observed in the control CZ medium (Fig. 3A and B). By comparison, velutinous or slightly floccose colonies with abundant sporulation and sparse white mycelium were observed when glyphosate was used to replace the phosphorus source (Fig. 3C). The color of conidia was mostly golden yellow unlike the color of conidia on the CZC medium. On the CZN medium, the colony morphology was completely different from that obtained on the other tested media. Plane and thin colonies with sparse sporulation were observed (Fig. 3D).
 (A), Czapek Dox medium without sucrose and supplied with glyphosate as the only carbon source CZC (B), Czapek Dox medium without K2HPO4 or NaNO3 and supplied with glyphosate as the only phosphorus CZP (C) and nitrogen source CZN (D), at 10 days of incubation.")
Morphological characteristics of colonies from Aspergillus section Flavi strains in control Czapek Dox medium (CZ) (A), Czapek Dox medium without sucrose and supplied with glyphosate as the only carbon source CZC (B), Czapek Dox medium without K2HPO4 or NaNO3 and supplied with glyphosate as the only phosphorus CZP (C) and nitrogen source CZN (D), at 10 days of incubation.
Similar macroscopic changes in colony morphology were observed in A. niger strains. No changes in the color of conidia were observed among the four assayed media. Colonies were plane when the herbicide was supplied as sole source of carbon. Abundant white mycelium and sparse sporulation in the center of the colonies were observed (Fig. 4B). Similarly to Aspergillus section Flavi, velutinous or slightly floccose colonies with abundant production of black conidia were observed on the CZP medium (Fig. 4C). Finally, the A. niger strains produced colonies with scarce presence of mycelial growth and conidial heads on the CZN medium (Fig. 4D).
 (A), Czapek Dox medium without sucrose and supplied with glyphosate as the only carbon source CZC (B), Czapek Dox medium without K2HPO4 or NaNO3 and supplied with glyphosate as the only phosphorus CZP (C) and nitrogen source CZN (D), at 10 days of incubation.")
Morphological characteristics of colonies from A. niger aggregate strains in control Czapek Dox medium (CZ) (A), Czapek Dox medium without sucrose and supplied with glyphosate as the only carbon source CZC (B), Czapek Dox medium without K2HPO4 or NaNO3 and supplied with glyphosate as the only phosphorus CZP (C) and nitrogen source CZN (D), at 10 days of incubation.
The variation of glyphosate levels in soil depends directly on the application rates as well as on the periodicity of rainfall. In relation to the soils from Argentina, Peruzzo et al.29 determined that levels of glyphosate in sediments and soils in a soybean cultivation area ranged from 0.5 to 5.0mg/kg whereas Botero et al.9 detected glyphosate and AMPA in a concentration of approximately 1mg/kg. Furthermore, these compounds were found in levels higher than 20mg/kg in seven samples of soybeans, from a total of eleven samples analyzed40. A new regulation raised the permitted glyphosate levels in oilseed crops, including soybean, from 20 to 40ppm17.
Interestingly, we found that four nontoxigenic Aspergillus section Flavi and two A. niger aggregate strains are able to grow consistently in a synthetic medium supplied with 10, 1.0 and 1.5mM glyphosate as sole carbon, phosphorus or nitrogen source, respectively. The length of the lag phase and the mycelial growth rate of the Aspergillus strains varied depending on the species and the media tested. The clear decrease of these parameters observed in the WA medium indicates that all the strains have a fast adaptation time to this medium lacking nutrients. The Aspergillus section Flavi strains need a longer adaptation time to the media supplied with glyphosate than to the control media. In contrast, the A. niger aggregate strains showed a shorter adaptation time in supplemented media, as compared to the control, except in the case of AN 384 in the medium supplied with glyphosate as source of carbon. The significant increase of the lag phase prior to growth observed under certain conditions such as Aspergillus section Flavi strains grown on CZN, CZP and CZC media and A. niger aggregate (strain AN 384) on CZC compared to the CZ medium suggests an induced nature of the degradation process.
In the literature, there are conflicting results regarding the impact of glyphosate on fungal growth and few data on its effect on Aspergillus strains. In a previous work, Carranza et al.11, reported lag phases values of 18–23h from nontoxigenic Aspergillus section Flavi strains grown on low nutrient status medium (soil-based medium) supplied with 5–20mM of glyphosate at −0.70MPa. However, the values obtained in this work in media supplied with glyphosate, except for strain AN 251 (lag phase 9h in CZN medium), are lower than those mentioned above. By comparison with the present results, the A. niger aggregate strains had previously shown longest lag phases (30–51h) when grown under the same conditions mentioned above12. These results show that several factors such as water availability, nutritional status of the medium and tested strains, among others, are involved in the length of the lag phase. This adaptation period is explained as the time necessary for the cells to synthesize the enzymes that are essential for growth when they have been introduced into a new environment. This phase or acclimation time is frequently observed between the time a chemical enters an environment and the time microbial degradation is evident2.
When studying the growth rate, we observed that the Aspergillus section Flavi strains showed a significant increase in this parameter in the treatment with glyphosate as nitrogen source. In addition, similar values were observed between the CZC and the CZP media. In the A. niger aggregate strains, the highest values depended on the strain and were found both in the CZN and the CZP media. The growth rate values obtained in this study with the Aspergillus section Flavi strains (around 4.5mm/day) are lower than those previously found with these fungi growing on soil-based medium supplied with 5–20mM of glyphosate at a higher water potential (−1.38). However, similar values were found at −2.78 and −7.06MPa11. With regard to the A. niger aggregate strains, lower growth rate values were observed in this study as compared to those previously found at −1.38 and −2.78MPa12.
Reddy et al.32 reported that two A. flavus strains isolated from agricultural soils had similar radial growth values on rich medium, control medium and medium supplied with 5 and 10mM of glyphosate after 14 days of incubation. Nevertheless, the radial growth of both strains was reduced by approximately 50 and 80% at 5 and 10mM of glyphosate, respectively, in a medium lacking nutrients. Although the conditions of that study were different from those in the present work, the results would be comparable even though no inhibition of growth was observed in the different media supplied with glyphosate and the response of the Aspergillus strains depended on the medium composition and the way the herbicide is used.
Hasan18 evaluated the capacity of Aspergillus spp. to use organophosphate pesticides in vitro and in soil. This author reported that the number of A. flavus colonies increased with 0.5mM of glyphosate used as the only source of carbon. A. niger and A. flavus, among other fungi, used and grew in the presence of 5 pesticides (glyphosate among them) as phosphorus source and showed more than a 50% increase in mass growth. In addition, they showed a production of phosphatase enzymes, which efficiently hydrolyzed the pesticides, suggesting that these species are important degraders of organophosphate compounds. Krzyśko-Łupicka et al.21 observed that glyphosate (at 10mM) as phosphorus source in the CZ medium supported exceptionally well the growth of three fungi including A. niger. However, their strains only grew on the CZ medium supplied with this pesticide as the sole source of phosphorus. Previous findings have suggested the ability of some fungi to biodegrade glyphosate and to use it as a source of phosphorus, whereas only a limited number of strains (Mucor spp., Rhizopus spp. and Fusarium spp.) would use the herbicide as a source of carbon13,20. Studying other fungi, Krzyśko-Łupicka and Sudol22 reported that Fusarium solani and Fusarium oxysporum strains do not use glyphosate as a source of carbon but as a source of phosphorus. In the presence of glyphosate (as the sole source of phosphorus) applied at concentrations of 1.0 and 1.5mM, the increase in the dry mass of the tested fungi was highly significant. Only one strain was capable of using glyphosate as the sole source of nitrogen. Similarly, Arfarita et al.4 showed that five genera of soil-borne fungi, among them Aspergillus sp., were able to grow consistently on a standard medium supplied with glyphosate. In addition, Scopulariopsis sp. and Verticillium sp. had a significant growth in CZ medium containing glyphosate as the sole source of phosphorus. In summary, most of the works reported the use of glyphosate as carbon or phosphorus source by fungal species. The results from the present work partially agree with those from previous studies due to the capacity of the Aspergillus strains to develop in culture media supplied with glyphosate not only as phosphorus or carbon source but also as a source of nitrogen. Other important aspect to consider in this study is that more than one isolate was used to represent each species. The inherent biological variation may be substantial among different individuals of the same species. Therefore, the effects may vary depending on the strains being studied.
Overall, the morphological changes in different culture media were similar in the two studied species. Velutinous or slightly floccose colonies with abundant sporulation were observed when glyphosate replaced the phosphorus source. By comparison, colonies presented sparse sporulation on CZC or CZN media and their aspect was completely different from that in other media. A consistent coloration of conidia was observed among the three media in the A. niger aggregate strains. In the Aspergillus section Flavi strains, golden yellow colonies were observed on the CZP medium unlike the typically olive green colonies exhibited by this species in the rest of the evaluated media, especially in the control CZ medium.
There is no information on the macroscopic changes of the Aspergillus species growing in the presence of glyphosate (commercial formulations). Tanney and Hutchison39, who studied other fungal species isolated from boreal forest soil, reported a complete or partial inhibition of colony pigmentation and sporulation when growing fungi in rich media with low glyphosate levels (≥1μg/ml). Correspondingly, Krzyśko-Łupicka and Sudol22 reported that Fusarium strains showed morphological changes in the presence of glyphosate applied as sole source of phosphorus in similar concentrations to those used in the present study (0.5–2.0mM).
Several studies confirmed a significant reduction of sporulation and subsequent spore germination in phytopathogenic fungi in the presence of glyphosate (commercial formulations) in vitro23,35,41. Depending on the persistence of these effects, sensitive species may undergo a decrease in fitness relative to their tolerant counterparts when exposed to glyphosate. The results here partially concur with those mentioned above since inhibition of sporulation in the Aspergillus strains was only observed when glyphosate was used as a source of carbon or nitrogen. Apparently, sporulation is related to the metabolism developed by the strains more than to the concentration of the herbicide since, in culture media with similar concentrations (CZN and CZP), the sporulation pattern was different. In support of this fact, Alvarenga da Silva et al.3 recently reported on the excellent growth of A. sydowii, Penicillium raistrickii and Trichoderma sp. on solid culture medium in the presence of the organophosphate pesticide profenofos. Thus, the observed phenotypic changes result from changes in the genetic apparatus. However, phenotypic changes were not persistent. After inoculation of fungi in the CZ full medium, all the strains showed the typical morphological features. Overall, the changes in the coloration of the mycelium are induced by the stress caused by the application of the herbicide. Given the complexity of these morphological changes, it is not possible to deepen this discussion.
Glyphosate, among other pesticides, plays an important role in the modern agriculture of several countries. Glyphosate is recognized as a cost-effective method for controlling pests, thus improving productivity of crops and food quality. However, the excessive and frequent application of this herbicide is related to a risk of accumulation of its residues in agricultural crops, which represents a potential hazard to human health. The nontoxigenic Aspergillus section Flavi and A. niger aggregate strains evaluated in this study showed a satisfactory growth performance with glyphosate in vitro, especially when used as sole source of phosphorus or nitrogen. Consequently, it is possible to suggest that these strains have potential for degrading glyphosate.
Further microcosm studies will allow to investigate the optimal environmental conditions for these strains to degrade glyphosate and to study the decomposition products formed. Despite the fact that several fungi that degrade organophosphorus compounds have been reported, it is important to consider that these fungal strains, which are native from Argentine agricultural soils, are adapted to the areas where glyphosate is frequently applied. This fact suggests their possible use in local agriculture.
Ethical disclosuresProtection of human and animal subjectsThe authors state that no human or animal experiments have been performed for this research.
Confidentiality of dataThe authors state that they have followed the protocols of their work center on the publication of patient data.
Right to privacy and informed consentThe authors state that no patient data appears in this article.
Conflict of interestThe authors declare that they have no conflicts of interest.
This work was supported by Agencia Nacional de Promoción Científica y Tecnológica (ANPCYT-PICT – 2482/10) and Secretaría de Ciencia y Técnica, Universidad Nacional de Río Cuarto (SECYT-UNRC – 18/C391). C.S. Carranza is a fellow from Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), C.L. Barberis, S.M. Chiacchiera and C.E. Magnoli are members of the research career of CONICET.
