





La fibrosis quística (FQ) es la enfermedad hereditaria autosómica recesiva más frecuente en la población de origen caucásico. Está producida por mutaciones en el gen que codifica el regulador de la conductancia transmembrana de FQ. Este defecto conduce, entre otras, a la alteración de las secreciones respiratorias, lo que determina una predisposición para la colonización-infección broncopulmonar crónica, causa principal de la elevada morbilidad y temprana mortalidad de estos pacientes. La colonización por Staphylococcus aureus y Haemophilus influenzae es frecuente en niños menores de 10 años, aunque Pseudomonas aeruginosa con morfotipo mucoide es, con diferencia, el microorganismo más relevante en el adulto y causa principal del deterioro broncopulmonar progresivo. Como consecuencia del tratamiento antimicrobiano repetido y la alteración pulmonar se favorece el desplazamiento de los patógenos habituales y se aíslan con mayor frecuencia bacilos gramnegativos no fermentadores, entre los que destacan Stenothrophomonas maltophilia, Achromobacter spp. y Burkholderia cepacia complex. Las connotaciones particulares del propio proceso patológico y de los microorganismos implicados hacen recomendable reconocer el seguimiento microbiológico de la colonización-infección broncopulmonar en el paciente con FQ como una entidad diagnóstica propia.
Cystic fibrosis (CF), a condition produced by mutations in the gene that encodes the cystic fibrosis transmembrane conductance regulator, is the most prevalent autosomal-recessive hereditary disease in caucasian populations. Among other repercussions, this defect leads to an alteration of respiratory secretions and determines a predisposition for chronic bronchopulmonary colonization-infection, which is the main driver of the high morbidity and early mortality of CF patients. Colonization by Staphylococcus aureus and Haemophilus influenzae is frequent in children younger than 10 years, but mucoid Pseudomonas aeruginosa is by far the most relevant pathogen in adults with CF and is responsible for the progressive bronchopulmonary deterioration. As a consequence of repeated, long-lasting antimicrobial treatments and deterioration of lung function, colonization by multidrug-resistant Gram-negative bacilli, such as Stenotrophomonas maltophilia, Achromobacter spp. and Burkholderia cepacia complex, is also frequent in adult CF patients. The special characteristics of the pathologic process and the microorganisms implicated in CF make it advisable to consider microbiological follow-up of chronic bronchopulmonary colonization-infection in these patients a specific diagnostic entity.
La fibrosis quística (FQ) es la enfermedad hereditaria autosómica recesiva más frecuente en la población de origen caucásico y la primera causa de afección pulmonar crónica en la infancia. Su frecuencia estimada oscila entre 1/2.500 y 1/5.000 recién nacidos vivos, lo que establece una frecuencia de 1 portador por cada 25-50 individuos en la población general, aunque se observan importantes diferencias dependientes de los grupos étnicos y regiones geográficas.
La FQ se produce como consecuencia de mutaciones en el gen que codifica el regulador de la conductancia transmembrana de fibrosis quística (CFTR), situado en el brazo largo del cromosoma 7. El CFTR actúa como canal de cloro y se encuentra en todos los tejidos exocrinos. La mutación más común es la deleción de la fenilalanina 508 (ΔF508), que se encuentra hasta en el 70% de los individuos con FQ. El defecto en el transporte del ión cloro causa que estos pacientes tengan un sudor característicamente salado y conduce a una deshidratación de las secreciones del tracto respiratorio, pancreáticas, hepáticas, intestinales y genitourinarias aumentando su viscosidad. Como consecuencia, las principales manifestaciones clínicas son respiratorias, gastrointestinales y genitourinarias1.
Las alteraciones digestivas fueron la causa de la temprana mortalidad de estos pacientes hasta mediados del siglo pasado, lo que determinó que hasta el 70% de los niños con FQ falleciera antes de cumplir el primer año de vida. Los tratamientos específicos para corregir las deficiencias digestivas en primera instancia, seguidos posteriormente de otros para combatir las infecciones respiratorias por Staphylococcus aureus, el primer patógeno relacionado con la FQ, determinaron un aumento notable de la esperanza de vida de los pacientes con FQ, situada ya en los 36,5 años según el último informe de la Fundación Americana para la Fibrosis Quistica2. A partir de la segunda mitad del siglo xx, en parte consecuencia de estos avances, empieza a cobrar relevancia la infección broncopulmonar crónica por Pseudomonas aeruginosa, que ya desde hace décadas se sitúa como la principal causa de las todavía elevadas morbilidad y mortalidad relacionadas con la FQ3.
Consideraciones clínicas y microbiológicas de la colonización-infección broncopulmonar crónica en la fibrosis quísticaPatogénesis de la colonización/infección broncopulmonarLa actividad mucociliar es una de las principales barreras del tracto respiratorio frente a los microorganismos y otros agentes extraños. El epitelio ciliado transporta la secreción mucosa por el árbol traqueobronquial arrastrando a su paso cualquier partícula o microorganismo que encuentre en su camino. Para que este mecanismo sea efectivo, el moco, que ocupa la parte central de la vía, debe deslizarse sobre la superficie serosa que envuelve los cilios de las células del epitelio bronquial. La alteración del CFTR da lugar a un aumento de la reabsorción de cloro y sodio que se acompaña de una reabsorción pasiva de agua y produce deshidratación de la superficie del epitelio ciliado respiratorio, que impide el correcto deslizamiento del moco a través del árbol traqueobronquial1,3. Esto conlleva un estancamiento del moco, el cual servirá de caldo de cultivo idóneo para diversos microorganismos. Otro factor patogénico añadido es la respuesta inflamatoria exagerada, causada por la propia infección y por la alteración presente en el epitelio bronquial, y que se manifiesta principalmente por un intenso infiltrado de neutrófilos, los cuales, mediante la secreción de proteasas, dañarían aún más el tejido bronquial. Además, el acúmulo de ADN, liberado principalmente por la lisis de los neutrófilos, incrementa la densidad y la viscosidad de las secreciones.
Además de la propia alteración cualitativa y cuantitativa de las secreciones mucosas y el deficiente aclaramiento mucociliar, hay varias hipótesis, no sin cierta controversia, que tratan de justificar la enorme predisposición de los pacientes con FQ para la infección broncopulmonar crónica por patógenos bacterianos determinados, particularmente por P. aeruginosa3,4. Entre ellas, cabría destacar especialmente tres: a) la alteración del CFTR conduce a la deshidratación de las secreciones respiratorias, con importante aumento de su osmolaridad, lo cual determina la inactivación de las betadefensinas, péptidos antibacterianos naturales que forman parte del sistema inmunitario innato; b) las células epiteliales de los pacientes con FQ son deficientes en la sialización de gangliósidos. Los asialogangliósidos actuarían como receptores para P. aeruginosa aumentando su adhesión al epitelio y, por lo tanto, su persistencia en las vías respiratorias, y c) el CFTR actúa como receptor para el LPS, de tal forma que su alteración determina una aclaramiento de P. aeruginosa por las células epiteliales entre 10 y 50 veces menor en los pacientes con FQ, lo que favorece su persistencia en las vías respiratorias.
Finalmente, varios estudios han demostrado que la concentración de hierro está aumentada en las secreciones respiratorias de los pacientes con FQ, lo cual parece favorecer la persistencia de la colonización por P. aeruginosa, según muestra un trabajo reciente. Asimismo, estudios recientes indican que la alteración del CFTR en los macrófagos alveolares reduce la acidificación de los fagolisosomas, lo cual, limita su capacidad bactericida y, por lo tanto, promueve la persistencia de la infección. De igual forma, las células del epitelio respiratorio de los pacientes con FQ son incapaces de secretar tiocianato, por lo que disminuye drásticamente su capacidad bactericida a través de la formación de moléculas de oxígeno reactivo.
Por lo tanto, debido a todos estos condicionantes, el paciente con FQ padecerá un cuadro de infección broncopulmonar crónica que se irá exacerbando a lo largo de su vida y que, a pesar de los avances en la antibioterapia y en el tratamiento con agentes mucolíticos y enzimas que rompen el ADN acumulado, sigue siendo, con diferencia, la causa principal de su peor calidad de vida y menor expectativa de supervivencia.
Cronoinfección broncopulmonar en la fibrosis quísticaEl análisis del patrón y la evolución temporal de la colonización en los pacientes con FQ ha permitido definir el concepto de cronoinfección por el que los pacientes sufrirían infecciones o colonizaciones siguiendo una secuencia más o menos establecida dependiente de la edad3,5,6. En los de menor edad, las infecciones por virus respiratorios y micoplasmas no son infrecuentes. Con posterioridad y aun en la primera década de la vida, es frecuente el aislamiento de Streptococcus pneumoniae y Haemophilus influenzae que serían rápidamente relegados a un segundo plano y sustituidos por S. aureus y posteriormente por P. aeruginosa. En la edad adulta, más del 80% de los pacientes están crónicamente colonizados por este microorganismo y en la mayoría de los casos se aísla en su morfotipo mucoso.
Como consecuencia del tratamiento antimicrobiano repetido en los pacientes adultos y del deterioro de la función pulmonar, se favorece el desplazamiento de los patógenos bacterianos habituales y se aíslan con mayor frecuencia bacilos gramnegativos no fermentadores entre los que destacan Stenotrophomonas maltophilia, Achromobacter spp. y Burkholderia cepacia. En el Hospital Ramón y Cajal, en el seguimiento de 81 pacientes con FQ durante 5 años, se observó una mayor incidencia de S. aureus en los primeros años de vida y un ulterior aumento de los pacientes colonizados con P. aeruginosa (fig. 1). Estos datos son muy similares a los del registro estadounidense de pacientes con FQ2. El aislamiento de las micobacterias atípicas en las secreciones respiratorias en pacientes adultos no es infrecuente. El tratamiento prolongado con antimicrobianos en la edad adulta también facilita el asentamiento de levaduras y hongos filamentosos, que complican aún más el patrón de colonización bronquial. Por último, el patrón de colonización bronquial no siempre es monomicrobiano y hasta en el 70% de los pacientes pueden coexistir diferentes patógenos. En más del 50% de ellos aparecen simultáneamente S. aureus y P. aeruginosa, solos o en asociación con con H. influenzae o S. pneumoniae.
Principales microorganismos implicados en la colonización-infección broncopulmonar crónicaPseudomonas aeruginosa por intervalos de edades en pacientes con fibrosis quística. Seguimiento de 5 años de 81 pacientes en la Unidad de Fibrosis Quística del Hospital Universitario Ramón y Cajal.")
Es con diferencia el microorganismo más frecuente en estos pacientes. Cerca del 60% de ellos está crónicamente colonizado por este microorganismo, que llegando al 80% en pacientes mayores de 18 años. La colonización-infección por P. aeruginosa se relaciona claramente con mayores morbilidad y mortalidad en el paciente con FQ. Se ha evidenciado un progresivo deterioro de la función pulmonar y una disminución de la supervivencia, y se ha comprobado que su adquisición en edades tempranas influye negativamente en el pronóstico de la enfermedad1. Los pacientes colonizados por P. aeruginosa durante los primeros 5 años de vida tienen un riesgo mayor de mortalidad (2,6 veces) que el de los pacientes con FQ no colonizados por este microorganismo. También se observan unos valores significativamente más bajos de FEV1 (volumen espiatorio forzado en el primer segundo), menor percentil de peso y aumento del número de hospitalizaciones. Los factores de riesgo para la colonización-infección temprana por P. aeruginosa incluyen la infección previa por S. aureus, el sexo femenino, la presencia homocigótica de la mutación ΔF508 y el contacto previo con pacientes adultos con FQ.
La adquisición de este patógeno se produce a partir de microorganismos presentes en el ambiente, aunque la transmisión cruzada entre pacientes no es del todo infrecuente, particularmente entre miembros de una misma familia. Asimismo, se han descrito brotes epidémicos de colonización-infección por P. aeruginosa en varias unidades de FQ7.
Inicialmente, la colonización del tracto respiratorio se produce por morfotipos no mucosos, generalmente sensibles a los antimicrobianos, y se presenta con una baja densidad bacteriana. Con posterioridad, y durante un período de tiempo variable, los cultivos de las muestras respiratorias pueden ser intermitentes. Una vez establecida la colonización-infección pulmonar crónica, generalmente por una única línea clonal de P. aeruginosa, resulta prácticamente imposible conseguir la erradicación.
El proceso de adaptación determinante de la persistencia (en la mayoría de los casos de por vida) de P. aeruginosa en las vías respiratorias de los pacientes con FQ incluye cambios tanto fisiológicos como genéticos8. Entre los primeros destaca la transición desde el estado de crecimiento planctónico (células libres suspendidas en medio acuoso) al de crecimiento en biopelículas o biofilms. Las biopelículas son estructuras supracelulares (comunidades multicelulares) complejas y bien organizadas espacial y funcionalmente que crecen sobre una superficie viva o inerte. La transición al crecimiento en forma de biopelículas depende de la acción de los sistemas de comunicación intercelular lasR-lasI y rhlR-rhlI, que son activados cuando la población alcanza una suficiente densidad, y por ello se denominan sistemas sensores de quorum (quorum sensing). El crecimiento en forma de biopelículas confiere al microorganismo una notable resistencia tanto a los tratamientos antibióticos como a la propia respuesta inmunitaria del paciente, que favorece su persistencia.
Además del crecimiento en forma de biopelículas, el desarrollo de la infección crónica por P. aeruginosa se materializa a través de un intenso proceso de adaptación genética, que será determinante para su resistencia a las condiciones ambientales (incluidos nuevamente la respuesta inmunitaria y los antibióticos). Fruto de esta intensa adaptación emergen gran cantidad de variantes fenotípicas características de la infección crónica, entre las que destacan la hiperproducción constitutiva de alginato (morfotipo mucoso) o los variantes de lento crecimiento (colonias enanas o small colony variants, SCV). Otras variantes fenotípicas que parecen favorecer la persistencia en las vías respiratorias son los mutantes aflagelados o con modificaciones del LPS. Se ha comprobado que la conversión al morfotipo mucoide se correlaciona con producción de anticuerpos y se acompaña de cambios importantes en los parámetros pulmonares, lo que conlleva una mayor mortalidad9. Asimismo, el incremento en el número de variantes morfológicas se ha correlacionado con el progresivo deterioro de la función pulmonar. Recientemente se ha comprobado que el intenso proceso de adaptación genética durante la colonización crónica favorece la persistencia del microorganismo en las vías respiratorias10.
Estas características, a pesar de no formar parte de los mecanismos específicos de resistencia a antibióticos, establecen una línea base de resistencia in vivo propicia para el desarrollo y la selección de variantes con resistencia a múltiples antibióticos (cepas multirresistentes), como consecuencia del uso prolongado de antibióticos. Este hecho se ve facilitado debido a que P. aeruginosa tiene una extraordinaria capacidad de desarrollar resistencia mediante mutaciones cromosómicas a prácticamente todos los antibióticos utilizados, incluidos los betalactámicos, los aminoglucósidos y las fluoroquinolonas. Finalmente, este desalentador panorama se agrava por la elevada prevalencia en los pacientes con FQ de cepas hipermutadoras, que presentan una frecuencia de mutación espontánea (para cualquier gen, incluyendo los implicados en la resistencia a antibióticos u otras mutaciones adaptativas, como las anteriormente mencionadas) hasta 1.000 veces mayor de lo normal. Entre el 30 y el 60% de los pacientes con FQ están colonizados por cepas hipermutadoras, según varios estudios, hecho extremadamente infrecuente (<1%) en pacientes con infecciones agudas11. La base molecular del fenotipo hipermutador es, en la mayoría de los casos, la deficiencia de alguno de los genes que forman parte del sistema de reparación de emparejamientos erróneos de ADN. Las cepas hipermutadoras aisladas de los pacientes con FQ son mucho más resistentes a los antibióticos que las no hipermutadoras; de hecho, la mayor prevalencia de cepas resistentes en los pacientes con FQ que en otros procesos se debe, en gran parte, a la contribución de las cepas hipermutadoras a las cifras de resistencia. Asimismo, mediante estudios in vitro e in vivo se ha demostrado que las cepas hipermutadoras desarrollan resistencia en pocas horas a prácticamente todos los antibióticos antipseudomónicos cuando se utilizan en monoterapia12. Recientemente se ha documentado que la hipermutación y su relación con la resistencia a los antibióticos no son exclusivas de la FQ, sino que también son muy frecuentes en el contexto de otras enfermedades respiratorias crónicas como bronquiectasias o en la enfermedad pulmonar obstructiva crónica (EPOC)13.
Staphylococcus aureusFue el primer microorganismo reconocido como causante de infección pulmonar crónica en pacientes con FQ. En la era preantibiótica fue uno de los principales implicados en la elevada y temprana mortalidad de estos pacientes. La terapia antiestafilocócica ha conseguido en las últimas décadas una notable reducción de la morbilidad y la mortalidad por este microorganismo, pero aún sigue siendo, después de P. aeruginosa, uno de los principales patógenos implicados en la infección broncopulmonar, especialmente en niños menores de 10 años. Los datos de 2005 del registro de pacientes de la Fundación Americana de FQ muestran que, de forma general, el 52% de los pacientes están colonizados por S. aureus2. Además, es el principal patógeno aislado durante los primeros 10 años de vida y alcanza su mayor prevalencia (>60%) en el grupo de 6-10 años de edad.
La persistencia de S. aureus depende fundamentalmente de su capacidad de adhesión al epitelio respiratorio y para evadir la respuesta inmunitaria una vez producida la colonización. Investigaciones recientes indican que S. aureus escapa más eficientemente de los fagosomas de las células del epitelio bronquial deficientes en el CFTR14. Además, se desarrolla adecuadamente en medios con alta osmolaridad, situación que se produce en las vías respiratorias del paciente con FQ. S. aureus puede persistir en las vías respiratorias de los pacientes con FQ sin producir alteraciones aparentes. No obstante, se ha apuntado que causa una lesión inicial con escasa repercusión funcional que predispondría a la colonización posterior por P. aeruginosa. Si bien el tratamiento profiláctico con betalactámicos ha mejorado notablemente el pronóstico de los niños con FQ, hay cierta controversia en torno al tratamiento indiscriminado, ya que un tratamiento prolongado podría llevar a la sustitución de S. aureus por P. aeruginosa y, por lo tanto, a adelantar el proceso evolutivo de la infección crónica y el deterioro pulmonar. Por otro lado, si bien la resistencia a meticilina no ha sido tradicionalmente un problema importante en los pacientes con FQ, la situación parece estar cambiando. Los datos de 2005 de la Fundación Americana muestran que actualmente hasta el 17,2% de los pacientes con FQ están colonizados por S. aureus resistente a meticilina (SARM). Recientemente se ha documentado la infección respiratoria por clones de SARM de origen extrahospitalario productores de la leucocidina de Panton-Valentine (LPV) en pacientes con FQ de Estados Unidos, situación que podría modificar la patogenia de la infección-colonización por este microorganismo. La situación actual en España de la resistencia a la meticilina en el contexto de FQ ha sido poco estudiada, pero puesto que se han detectado recientemente los primeros casos de SARM de origen extrahospitalario, podría seguir el mismo camino descrito en América y, por lo tanto, será necesario mantener una vigilancia activa de este fenómeno en los próximos años.
Otro fenómeno preocupante desde el punto de vista de la resistencia a antibióticos es la alta prevalencia (el 14% según un estudio publicado) de cepas hipermutadoras de S. aureus en los pacientes con FQ, al igual que ocurre para P. aeruginosa15. Si bien la hipermutación aparentemente no contribuye al desarrollo de resistencia a la meticilina, puesto que ésta se produce por la adquisición de determinantes exógenos de resistencia y no por mutación, sí contribuye notablemente al desarrollo de resistencia a antibióticos como los macrólidos; facilitando la selección de mutaciones en genes ribosómicos. En cualquier caso, la resistencia a macrólidos es cada vez más frecuente en las cepas de S. aureus de pacientes con FQ, al menos en parte debido al uso extendido de la terapia de mantenimiento con azitromicina. Asimismo, el tratamiento profiláctico prolongado con cotrimoxazol se ha relacionado con la selección de cepas dependientes de timidina por inactivación del gen thyA que codifica para la timidilato sintetasa. Estas cepas, además de ser resistentes al cotrimoxazol, se caracterizan por producir colonias con morfologías anómalas (SCV) que incluso muchas veces no crecen en medios convencionales y pueden pasar inadvertidas16. Estudios recientes demuestran que la coinfección con P. aeruginosa (situación que ocurre de forma frecuente) también determina la selección de variantes SCV de S. aureus. Este hecho, además de plantear dificultades diagnósticas, conlleva una mayor resistencia a los antibióticos y una mayor capacidad de persistencia en las vías respiratorias de estos pacientes.
Haemophilus influenzaeAunque su frecuencia de aislamiento varía según el protocolo microbiológico aplicado, en la mayoría de los trabajos es el tercer microorganismo más frecuentemente aislado en los pacientes con FQ después de P. aeruginosa y S. aureus. Su frecuencia aumenta si se emplean medios selectivos o incubación en atmósfera con anaerobiosis5,17.
H. influenzae puede colonizar hasta al 30% de los pacientes con FQ. Su incidencia es mayor en los niños de menor edad, si bien también puede aislarse en los pacientes adultos. No hay evidencias claras sobre su papel como factor primario en el deterioro de la función pulmonar y su efecto patogénico estaría en relación con la carga bacteriana elevada y la respuesta inflamatoria generada. Aunque no es habitual, hay pacientes con colonización crónica, por nuevas cepas o por cepas persistentes, que siguen un patrón similar al que presentan los bronquíticos crónicos. En los pacientes con FQ se ha encontrado relación con las exacerbaciones durante el periodo de colonización crónica que suelen ser generalmente breves, remiten con el tratamiento antimicrobiano adecuado e incluso puede erradicarse hasta en el 70% de los casos de las secreciones respiratorias. Cuando esto ocurre, se reducen los anticuerpos contra este microorganismo y se evidencia una mejoría clínica con desaparición o reducción de los síntomas.
Las cepas de H. influenzae que se aíslan en el paciente con FQ son típicamente no capsuladas, característica común con los encontrados en otras enfermedades respiratorias que cursan con cronicidad18,19. Por ello la vacuna conjugada contra H. influenzae tipo b no tendría utilidad en la prevención de la colonización en el paciente con FQ. El biotipo más frecuentemente aislado en estos pacientes es el I, que es uno de los que conllevan una mayor virulencia.
Aunque no hay evidencia clínica, hay acuerdo generalizado del beneficio del tratamiento antimicrobiano en los pacientes con FQ colonizados por este patógeno. Las pautas aplicadas no difieren de las utilizadas en las infecciones respiratorias por este agente en otro tipo de pacientes. Entre el 20 y el 40% de los aislados de H. influenzae son resistentes a la amoxicilina debido a la producción de betalactamasas. Un número relativamente importante de las cepas de H. influenzae en los pacientes con FQ es resistente a amoxicilina sin producir betalactamasa. Este fenotipo, denominado BLNAR (beta-lactamase-negative ampicillin-resistant), es debido a alteraciones en las PBP que confieren también pérdida de sensibilidad a la asociación de amoxicilina-clavulánico y cefalosporias orales. Su incidencia en un amplio estudio realizado con pacientes seguidos en la Unidad de FQ del Hospital Universitario Ramón y Cajal fue algo mayor del 5%, pero podrían estar aumentando en los últimos años19.
Se ha descrito la persistencia prolongada en los pacientes con FQ, incluso de años, de cepas de H. influenzae multirresistentes a pesar del tratamiento adecuado con antimicrobianos. Este perfil de multirresistencia parece estar relacionado, al igual que en P. aeruginosa, con cepas hipermutadoras. En España, se han descrito casos de pacientes persistentemente colonizados por H. influenzae resistentes a las fluoroquinolonas. Este hallazgo es excepcional y la selección del mecanismo de resistencia implicado en ello, alteraciones en gyrA y parC, se relaciona con la administración previa de fluoroquinolonas y el fenotipo hipermutador19,20.
Streptococcus pneumoniaeEn la mayoría de las series, su incidencia no supera el 20%, y es mayor en las etapas iniciales de la enfermedad, en que puede llegar hasta el 50%. En el Hospital Universitario Ramón y Cajal durante el seguimiento de los pacientes atendidos en su Unidad de FQ entre 1995 y 2003, se encontró una incidencia general anual media del 5,5%; todos los pacientes en los que se aisló tenían menos de 12 años edad21. En el 35% de los casos, su aislamiento tenía relación con un episodio de exacerbación, aunque sólo en el 27% de éstos se identificó como único patógeno durante estos episodios. La utilización de medios selectivos, por ejemplo agar sangre suplementado con ácido nalidíxico y colistina, no parece incrementar el porcentaje de aislamientos de S. pneumoniae en los pacientes con FQ.
En cuanto al aspecto de la patogenia, S. pneumoniae se comportaría como S. aureus, que es capaz de adherirse a la superficie mucosa, lo que favorece las infecciones broncopulmonares crónicas. Otro posible factor de virulencia, común con P. aeruginosa y S. aureus, es su capacidad para elaborar productos extracelulares que estimulan la secreción mucosa y contribuyen a una peor evolución de las exacerbaciones. Recientemente se ha demostrado una mayor facilidad de los aislados de S. pneumoniae obtenidos de los pacientes con FQ para formar biopelículas en comparación con los obtenidos de enfermedades invasivas y que también explicaría su posible facilidad para permanecer en el árbol bronquial22. Al igual que H. influenzae, puede producirse una persistencia de clones específicos en muchos casos multirresistentes, que presentan tasas de mutación mayores que la de los aislados que no proceden de estos pacientes. La detección, en estos pacientes, de serotipos similares a los que cubre la vacuna heptavalente contra S. pneumoniae justificaría su utilización y podría reducir el estado de portador por serotipos multirresistentes.
Burkholderia cepacia complexSe ha comprobado que los organismos identificados como B. cepacia forman un grupo muy heterogéneo y constituyen un complejo de especies fenotípicamente similares23. Hay al menos 9 variedades genómicas dentro del complejo: B. cepacia (variedad genómica I), B. multivorans (variedad genómica II), B. cenocepacia (variedad genómica III), B. stabilis (variedad genómica IV), B. vietnamiensis (variedad genómica V), B. dolosa (variedad genómica VI), B. ambifaria (variedad genómica VII), B. anthina (variedad genómica VIII) y B. pyrrocina (variedad genómica IX) y otras 15 especies dentro del género Burkholderia24. La variedad genómica I es un patógeno de plantas y contiene la especie tipo. La mayoría de los aislamientos de pacientes con FQ se incluyen dentro de B. cenocepacia y B. multivorans.
B. cepacia apareció hace más de 25 años como un importante patógeno oportunista en estos pacientes. Con frecuencia, la infección pulmonar producida por esta bacteria es crónica, difícil de tratar con antimicrobianos por su multirresistencia y conlleva importantes morbilidad y mortalidad. En estos pacientes, se ha asociado con lo que se ha denominado síndrome cepacia, que se caracteriza por fiebre alta, bacteriemia, bronconeumonía, deterioro pulmonar rápido y muerte en más del 60% de los casos. La transmisión entre pacientes se puede producir por contacto directo o indirecto con secreciones de pacientes infectados y se facilita por contacto prolongado entre pacientes con FQ, por compartir equipos, por contacto social, tanto en el medio hospitalario como en reuniones, campamentos, etc.24. En algunos centros se excluye a los pacientes colonizados con B. cepacia de los programas de trasplante pulmonar, debido al mal pronóstico23,24. Sin embargo, para otros autores sería necesario valorar la infección con diferentes especies dentro del complejo de forma individualizada.
Los estudios epidemiológicos indican que las diferentes variedades genómicas son diversas en cuanto a su frecuencia de transmisión y patogenia24. B. cenocepacia es frecuente en suelo agrícola, pero no en sitios urbanos, y hasta ahora no está claramente demostrado que los pacientes se contagien de fuentes ambientales. Conlleva una elevada transmisión y mal pronóstico y la transmisión entre pacientes se relaciona con uno o dos marcadores genéticos: gen cblA («cable Pili») y BCESM (B. cepacia epidemic strain marker), aunque no se ha encontrado en las cepas de todos los brotes. B. gladioli es un patógeno de plantas aislado tradicionalmente del gladiolo y el arroz. Se ha aislado de muestras de pacientes con FQ, y se ha identificado por su capacidad para crecer en medios selectivos para el aislamiento de B. cepacia. B. gladioli y B. cepacia complex son genotípicamente diferentes pero fenotípicamente muy similares y es difícil diferenciarlas mediante técnicas convencionales25.
Stenotrophomonas maltophilia, Achromobacter spp. y otros bacilos gramnegativos no fermentadoresS. maltophilia y Achromobacter xylosoxidans se encuentran más frecuentemente que B. cepacia complex en pacientes con FQ y enfermedad pulmonar avanzada, pero generalmente son menos virulentos.
S. maltophilia es un bacilo gramnegativo no fermentador, oxidasa negativo y resistente a la mayoría de los antimicrobianos. Hay factores que predisponen a la colonización por este microorganismo, como el uso prolongado de antimicrobianos. Sin embargo, parece que no predispone la hospitalización previa o el contacto con personas colonizadas, aunque sí se ha descrito colonización con la misma cepa en diferentes pacientes24. S. maltophilia suele producir una colonización transitoria, aunque en alrededor del 10% de los casos se ha documentado colonización crónica26. En un estudio, se observó una peor supervivencia a los 5 años de los pacientes con S. maltophilia; sin embargo, en otros no se evidencia un deterioro clínico relacionado con esta bacteria25. La prevalencia de colonización en pacientes con FQ ha aumentado en los últimos años, aunque varía de unos centros a otros, en parte, por el uso de diferentes medios selectivos o por los métodos de identificación que pueden dar lugar a identificaciones erróneas24.
A. xylosoxidans es también un bacilo gramnegativo que puede confundirse con P. aeruginosa no pigmentada o con B. cepacia25. No hay estudios que examinen el efecto de esta bacteria en la función pulmonar y en la mortalidad. En un estudio, se ha relacionado con exacerbaciones pulmonares en FQ, pero los pacientes estaban colonizados también por P. aeruginosa. No parece que la colonización a largo plazo produzca un deterioro claro del estado clínico. Se han descrito casos de pacientes colonizados con la misma cepa en el mismo centro, y aunque no se encontró la fuente, estuvieron hospitalizados al mismo tiempo24.
Ralstonia, género establecido en 1995, agrupa diferentes especies patógenas en humanos: R. pickettii, R. paucula, R. gilardii y R. mannitolilytica25. Son bacilos gramnegativos no fermentadores, oxidasa positivos, y se consideran patógenos oportunistas que pueden producir infecciones graves en pacientes inmunodeficientes. Estos organismos pueden crecer en los medios selectivos para B. cepacia, y con los métodos fenotípicos convencionales se pueden identificar de forma errónea17.
Pandoraea es un bacilo gramnegativo no fermentador que se ha aislado de muestras ambientales, muestras clínicas y recientemente en muestras de pacientes con FQ, que es capaz de crecer en medios selectivos para B. cepacia. Este género, descrito en 2000, contiene 5 especies: P. apista, P. sputorum, P. norimbergensis, P. pnomenusa y P. pulmonicola y otras 4 genoespecies todavía sin nombre (variedad genómica 1 a 4). El género más cercano filogenéticamente es Burkholderia y comparte características fenotípicas con Ralstonia. Parece que son potencialmente patógenos en pacientes con FQ y se ha descrito transmisión entre pacientes. Clínicamente es más virulenta que Ralstonia spp. o B. gladioli. La identificación fenotípica no es fiable y requiere técnicas moleculares17. Es resistente a muchos antimicrobianos, lo que limita las opciones terapéuticas.
Inquilinus sp. es un nuevo género descrito en 2002, al realizar estudios taxonómicos de las bacterias gramnegativas no habituales obtenidas de secreciones respiratorias de pacientes con FQ. Es un bacilo gramnegativo no fermentador que pueden crecer en medios selectivos para B. cepacia, pero puede no crecer en MacConkey17. En este género se incluye la especie I. limosus más otra aún sin nombre. Puede tener aspecto mucoide y es resistente a colistina y a todos los betalactámicos, excepto imipenem. No hay datos suficientes para conocer el significado clínico de estas bacterias en estos pacientes.
En pacientes con FQ se han aislado, además, otras bacterias gramnegativas, como Comamonas acidovorans, Comamonas testosteroni, Acinetobacter calcoaceticus o Chryseobacterium indologenes.
EnterobacteriaceaeLas enterobacterias se pueden aislar del tracto respiratorio de pacientes con FQ, aunque generalmente son colonizaciones transitorias y no están relacionadas con enfermedad grave17. Se pueden encontrar en el 1–4% de los pacientes, y en el 10% de los casos de exacerbaciones agudas. Se aíslan con mayor frecuencia en niños de 0 a 5 años, generalmente antes de que se produzca una colonización crónica por P. aeruginosa. Entre las enterobacterias, Escherichia coli es la que se aísla con mayor frecuencia, seguido de Klebsiella spp., Enterobacter spp., Citrobacter spp. y Proteus spp. Su detección puede ser compleja cuando está presente P. aeruginosa por competencia durante el crecimiento. Aunque no es muy frecuente, las enterobacterias pueden producir infecciones crónicas, desarrollando fenotipos altamente mucoides, similares a los de P. aeruginosa, con la que pueden llegar a confundirse a simple vista si no se realiza la prueba de la oxidasa. Al igual que con P. aeruginosa, la infección crónica conlleva el desarrollo de multirresistencia. Ocasionalmente, se ha descrito también la presencia de cepas productoras de betalactamasas de espectro extendido, lo cual dificulta el control de la colonización por estos microorganismos.
Mycobacterium spp.Los pacientes con FQ tienen un riesgo mayor de sufrir colonización-infección respiratoria por micobacterias no tuberculosas (MNT), probablemente debido a bronquiectasias e infecciones crónicas y recurrentes del pulmón3. A diferencia de lo que ocurre en el caso de la tuberculosis, el contagio se produce muy raramente por contacto persona a persona y el tipificado de las cepas aisladas en los pacientes ha demostrado la ausencia de transmisión cruzada entre ellos. Las MNT tienen una distribución amplia en la naturaleza y se encuentran principalmente en el agua, el suelo, las plantas y los animales. El aumento aparente en el aislamiento de MNT en estos pacientes probablemente esté relacionado con su mayor supervivencia y la mejora en los métodos de detección e identificación de las MNT27,28.
En series publicadas, la prevalencia varía ampliamente, según los centros, del 2 al 28%, si bien la media se situaría entre el 5 y el 15%27–29. Las prevalencias más bajas podrían deberse a series basadas en un solo cultivo, a no utilizar técnicas de descontaminación adecuadas para evitar el sobrecrecimiento de otras bacterias o la poca calidad de las muestras, sobre todo en pacientes pediátricos con dificultades para expectorar.
Las especies que se aíslan más frecuentemente son M. avium complex (MAC), M. avium con mayor frecuencia que M. intracellulare, seguida de M. abscessus. La edad de los pacientes parece influir en la frecuencia y las especies de MNT aisladas. En menores de 10–12 años la prevalencia sería del 4–5% y la más frecuente, M. abcessus, junto con otras MNT de crecimiento rápido como M. chelonae y M. fortuitum28. La prevalencia va aumentando con la edad, más del 15% en mayores de 15 años, lo que puede ser un reflejo de la cada vez mayor longevidad de los pacientes con FQ. M. abscessus se aísla en un porcentaje similar (5%) a cualquier edad, mientras que MAC se aísla fundamentalmente en mayores de 10–15 años. Otras MNT de crecimiento lento aisladas son M. gordonae y más raramente M. kansasii. En nuestro país no se ha publicado ningún estudio multicéntrico, pero sí se han notificado algunas series con frecuencia de aislamiento variable, del 4 al 25%29.
Las bacterias de crecimiento rápido en el esputo de los pacientes con FQ pueden ser una causa de que la prevalencia real de MNT esté subestimada. La tasa de cultivos contaminados, en el caso de pacientes con FQ (24–45%), contrasta claramente con la general (2,5–3%) del Laboratorio de Micobacterias. Principalmente P. aeruginosa, que se encuentra en el tracto respiratorio de más del 80% de estos pacientes y alrededor de un tercio de los aislados presentan fenotipo mucoso y sobreviven al tratamiento de descontaminación sistemático con N-acetil-L-cisteína+2% NaOH. La inoculación simultánea en medios sólidos, preferentemente Lowestein-Jensen, y medios líquidos mejora el número de micobacterias recuperadas y disminuye el de muestras finalmente informadas como contaminadas hasta cerca del 17%. Para eliminar eficazmente P. aeruginosa, el procesamiento previo de las muestras respiratorias de pacientes con FQ debería incluir tratamiento con ácido oxálico al 5% en un segundo paso, después de la descontaminación con N-acetil-L-cisteína+2% NaOH. Este tratamiento más agresivo reduce eficazmente la contaminación por bacterias de rápido crecimiento hasta un 6–8%, pero puede inactivar entre un 25 y un 30% de las micobacterias, sobre todo en muestras con una carga bacteriana baja. Para minimizar este efecto, se ha propuesto realizar la descontaminación en dos etapas con N-acetil-L-cisteína+2% NaOH y el ácido oxálico al 5% sólo del cultivo en medio líquido cuando resulte contaminado. De esta forma se puede llegar a obtener hasta un 25% de mejora en la sensibilidad.
Recientemente, se ha descrito la utilización de un método de descontaminación con cicloheximida que mejora el aislamiento de MNT, principalmente M. abscessus. Este método es más sencillo y rápido pero tiene el inconveniente de que la muestra descontaminada no puede inocularse a los medios líquidos automatizados. Esto se debe a que la adición de lecitina al medio líquido, necesaria para neutralizar la acción de la cicloheximida, da lugar a fluorescencia inespecífica que impide la lectura automática de los frascos.
La distinción entre infección pulmonar activa y colonización no es fácil en el paciente con FQ. Las recomendaciones publicadas por la American Thoracic Society (ATS) para el diagnóstico de infección pulmonar por MNT incluyen criterios bacteriológicos, clínicos y radiológicos; los dos últimos son muy difíciles de establecer. La definición de infección requiere un mínimo de tres esputos o lavados bronquiales positivos o 2 cultivos positivos con al menos una tinción positiva en los 12 meses previos. Si se obtiene un cultivo positivo para MNT, es obligatorio realizar cultivos seriados para comprobar si se aísla la misma especie de MNT y, en el caso de que sean negativos, prolongarlos al menos durante 1 año para descartar la infección. En bastantes pacientes la colonización es ocasional ya que los cultivos repetidos son negativos, sobre todo en el caso de M. avium. La tinción directa es de gran importancia, ya que los pacientes que presentan tinciones positivas cumplen criterios bacteriológicos con mayor frecuencia y se ha apuntado que podrían ser indicativas de infección activa. Asimismo, el aislamiento repetido de MNT se asocia significativamente con enfermedad pulmonar granulomatosa diagnosticada por necropsia en pacientes con FQ. Los pacientes con aislamiento de M. abscessus cumplen los criterios bacteriológicos con mayor frecuencia que los pacientes con cultivo positivo del grupo MAC. En aproximadamente dos tercios de los pacientes con M. abscessus se observan alteraciones en la tomografía computarizada y se han descrito con mayor frecuencia casos de enfermedad invasiva mortal. M. abscessus, además, puede ser la causa de infección diseminada tras el trasplante pulmonar y la posterior terapia inmunosupresora.
La identificación de las especies es esencial y puede requerir técnicas genéticas. Dentro del grupo MAC se deben utilizar medios de identificación que diferencien las especies M. avium de M. intracellulare. Las técnicas utilizadas son la hibridación con sondas DNA-RNA (AccuProbe), la amplificación de secuencias genéticas seguidas de hibridación (INNO-LIPA), restricción (PCR-RFLP del gen hsp65) o secuenciación genética (PCR+ secuenciación de la subunidad 16S del ARNr o del gen hsp65). M. abcessus es la más patógena y resistente de las micobacterias de crecimiento rápido. Es esencial diferenciarla de M. chelonae ya que el tratamiento en las infecciones por M. abcessus es especialmente difícil y puede requerir cirugía en las infecciones localizadas si la función pulmonar es adecuada. Pueden diferenciarse fácilmente por la tolerancia al cloruro sódico o la utilización del citrato, pero puede ser necesaria la amplificación seguida de restricción del gen hsp65 o secuenciación genética del 16S del ARNr o del gen hsp65.
El estudio de la sensibilidad se recomienda sólo en el caso de aislamientos que se consideren clínicamente significativos. En los aislamientos de MAC el tratamiento antibiótico está protocolizado, pero puede ser recomendable el estudio de la sensibilidad a los macrólidos (claritromicina). En el caso de M. abcessus, no hay un consenso sobre el tratamiento a utilizar, ya que esta micobacteria es especialmente resistente a numerosos antibióticos y la elección del tratamiento se debe guiar por el valor de las CMI en medio líquido o, dada la complejidad de estas técnicas, por los resultados obtenidos por la técnica de E-test.
Con respecto a la infección por M. tuberculosis, es rara en pacientes con FQ y se encuentran muy pocas referencias en la literatura por lo que es difícil conocer su incidencia. En las series que recogen este dato el porcentaje de M. tuberculosis es mucho menor (0–2%) frente al aislamiento de MNT (4–25%). A diferencia de las MNT que se encuentran en el medio ambiente, M. tuberculosis se transmite persona a persona y es una seria complicación para el paciente con FQ, sobre todo si se trata de aislados multirresistentes, por lo que es recomendable descartarla rápidamente mediante técnicas de detección e identificación genéticas directas cuando la tinción de la muestra sea positiva.
Aspergillus y hongos levaduriformesLa prevalencia de Aspergillus en muestras respiratorias de pacientes con FQ varía desde el 6 al 57% según la edad (suele aparecer durante la adolescencia), el lugar de residencia (afecta más a pacientes del medio rural) o las condiciones climáticas (mayor incidencia en zonas costeras). En aproximadamente el 75% persiste en cultivos sucesivos y su aislamiento normalmente representa colonización pulmonar. En el 2–10% de estos pacientes la principal complicación es la aspergilosis broncopulmonar alérgica (ABPA), una hipersensibilización a los antígenos de Aspergillus con estimulación de la respuesta inmunitaria celular y producción de anticuerpos específicos30. La prevalencia es baja en menores de 6 años y es más elevada en los pacientes en peores condiciones clínicas. Las principales manifestaciones de esta enfermedad son respiración sibilante, aparición de infiltrados pulmonares, bronquiectasias y fibrosis.
El diagnóstico clínico de ABPA es difícil en estos pacientes ya que las manifestaciones de la enfermedad no son fáciles de diferenciar de las exacerbaciones agudas que sufren. Tampoco lo es el diagnóstico microbiológico ya que el aislamiento de Aspergillus en el esputo no es suficiente para el diagnóstico de ABPA, pero sí se ha demostrado una correlación positiva entre anticuerpos específicos en suero y afectación de la función pulmonar. El tratamiento profiláctico con antibióticos predispone a la colonización por hongos, aunque raramente se han notificado casos de aspergiloma o aspergilosis invasiva en pacientes con FQ, excepto en trasplantados pulmonares. La presencia en el esputo, sobre todo, de la forma mucoide de P. aeruginosa puede inhibir el crecimiento de hongos, por lo que se recomienda el uso de medios de cultivo para hongos, como el agar Sabouraud glucosa, suplementados con antimicrobianos, como ciprofloxacino o amikacina. La especie mayoritaria es Aspergillus fumigatus, aunque otras especies también se han descrito en una proporción mucho menor. Scedosporium apiospermum (3–8,6%) sería el segundo en importancia en algunas series. A diferencia de Aspergillus, sus esporas se encuentran raramente en el ambiente y los mecanismos de transmisión y colonización crónica no están claros. Podría estar asociado también a episodios de ABPA, pero la presencia de reacción cruzada con determinados antígenos de A. fumigatus hace difícil demostrarlo en los pacientes que presentan anticuerpos frente a Aspergillus.
Las levaduras se aíslan con mayor frecuencia, el 75–78% de los pacientes con FQ, y Candida albicans es la especie mayoritaria. Se presenta en pacientes que reciben tratamiento prolongado con antibióticos o glucocorticoides y el origen es principalmente endógeno a partir de la propia flora del paciente. Se han notificado casos de colonización por la levadura melaninogénica Exophiala dermatitis, y su prevalencia es del 2 al 15% cuando se emplea un medio de cultivo específico (agar eritritol cloranfenicol, ECA) y se prolonga la incubación hasta 4 semanas. El riesgo de infección por esta levadura en pacientes con FQ está todavía por establecer.
Virus respiratorios y otros patógenosA pesar del papel central de las infecciones bacterianas, hay otros factores no bacterianos implicados en la progresión de la enfermedad pulmonar en los pacientes con FQ. Durante la última década se ha evaluado en estos pacientes el impacto de las infecciones por virus respiratorio sincitial (VRS), virus de la gripe, adenovirus, virus parainfluenza, rinovirus y, más recientemente, metapneumovirus, pero no por otros virus causales de infecciones respiratorias como coronavirus o bocavirus31. También se ha considerado la implicación de bacterias atípicas como Chlamydophila pneumoniae, Mycoplasma pneumoniae, Bordetella pertussis y Legionella pneumophila31. En general, en el 5–20% de los pacientes con exacerbaciones agudas se puede demostrar una etiología viral o por las bacterias señaladas, frente al 0–5% en pacientes en situación estable. La importancia de estas infecciones en los pacientes con FQ probablemente esté subestimada, ya que en la mayoría de los estudios el diagnóstico fue serológico y adolecen de la falta de técnicas directas y rápidas. Recientemente también se ha señalado a Pneumocystic jirovecci como un agente que podría tener un papel relevante en la FQ y cuya importancia estaría infravalorada.
La incidencia de infecciones virales en estos pacientes no es mayor que en la población general y en niños menores de 2 años incluso más baja, debido probablemente a una exposición menor. Sin embargo, la infección respiratoria puede ser mucho más grave con mayor afectación de vías bajas, mayor incidencia de exacerbaciones agudas, hospitalización y deterioro de la función pulmonar. Además, puede predisponer a la infección por bacterias al dañar el epitelio respiratorio y facilitar la adherencia bacteriana. Algunos autores han encontrado asociación con un riesgo mayor de infección por P. aeruginosa, o con una afectación mayor, indicada por el aumento en el título de anticuerpos anti-Pseudomonas, en los pacientes con infección crónica por este microorganismo. Un mayor número de infecciones virales al año se correlaciona con progresión de la enfermedad pulmonar con disminución del índice de Swachman, (FEV1) o la capacidad vital forzada (FVC) y se ha demostrado que un número significativo de episodios de exacerbación pulmonar son precedidos por infecciones respiratorias virales.
Tipos de muestras, recogida, transporte y conservaciónEl lavado broncoalveolar es la muestra considerada de referencia en el estudio microbiológico del paciente con FQ. Sin embargo, el esputo es la más utilizada, por su facilidad de obtención y buena correlación con el lavado broncoalveolar. En los pacientes en quienes no se puede obtener muestras de esputo o en los niños pequeños suele recurrirse a los aspirados bronquiales o a tomas retrofaríngeas. Otras muestras utilizadas son los broncoaspirados y cepillados bronquiales. La recogida de las muestras en estos pacientes debe seguir las consideraciones generales32.
Esputo. Su toma ha de evitar la contaminación con microbiota del tracto respiratorio superior, ha de recogerse en envases estériles y remitirse con la mayor celeridad para su estudio. Si el procesamiento no es inmediato, se recomienda mantener la muestra a 4°C, ya que aunque a temperatura ambiente la viabilidad de S. aureus y P. aeruginosa no se ve afectada en las primeras 24–48h, si bien puede afectar a los recuentos bacterianos. En el caso de H. infuenzae y S. pneumoniae los recuentos disminuyen drásticamente con el tiempo y los cultivos podrían ser falsamente negativos. Para evitarlo, puede congelarse mejor que mantenerse en refrigeración.
Lavado broncoalveolar. Está recomendado en los pacientes con escasa expectoración, con antibioterapia previa prolongada, en los casos particulares de sospecha de colonización por B. cepacia, cuando se vaya a aplicar técnicas de biología molecular o en el seguimiento de los pacientes sometidos a trasplante pulmonar.
Muestras retrofaríngeas. Su valor diagnóstico puede variar dependiendo de la edad del paciente y del patrón de colonización. En el caso de P. aeruginosa y en pacientes menores de 5 años el valor predictivo positivo es cercano al 95% y el valor predictivo negativo, del 40%, y son algo inferiores para S. aureus. Por el contrario, en pacientes jóvenes sin expectoración el valor predictivo positivo para P. aeruginosa es menor (83%) y algo mayor el valor predictivo negativo (70%); los valores correspondientes para S. aureus son del 91 y el 80%, respectivamente. Se ha demostrado que el valor diagnóstico aumenta cuando se incrementa el número de tomas orofaríngeas estudiadas.
Valoración y manejo de la muestras en el laboratorio de microbiologíaEn general, se recurre a la tinción de Gram para valorar la idoneidad del esputo para el cultivo. El cultivo debe realizarse con los esputos que presenten más de 25 leucocitos y menos de 25 células epiteliales por campo microscópico con bajo aumento. En los pacientes con FQ no se considera imprescindible su realización puesto que puede no ser suficientemente ilustrativa de los microorganismos presentes, ya que éstos pueden formar acúmulos en las secreciones (biopelículas), o de las células inflamatorias, que no siempre se distribuyen de forma homogénea. Se estima que con los criterios habituales de valoración de la tinción de Gram, hasta el 40% de las muestras de esputo de los pacientes con FQ serían inadecuadas para el cultivo y, sin embargo, ofrecen resultados valorables.
Las muestras respiratorias de los pacientes con FQ, y en particular los esputos, presentan una elevada consistencia y deben someterse a un proceso de homogeneización antes de proceder a su cultivo. Habitualmente se emplean agentes mucolíticos (N-acetilcisteína) o ditiotreitol. También se ha recomendado emplear una homogeneización mecánica (sonicación suave) o utilizar simplemente suero salino17.
Procesamiento de las muestras respiratorias para cultivo cuantitativoFinalidad y justificación del cultivo cuantitativoEl diagnóstico y el seguimiento microbiológico cualitativo y cuantitativo de la colonización-infección de las vías respiratorias son una herramienta clave para el manejo clínico de los pacientes con FQ. Según las últimas recomendaciones de la Fundación Americana de FQ, se deben obtener muestras respiratorias para estudio microbiológico en las siguientes premisas: a) al menos una cada 3 meses en pacientes clínicamente estables y sin exacerbaciones pulmonares; b) durante los cuadros de exacerbación pulmonar; c) en caso de cambio del estado clínico; d) en caso de hospitalización, y e) cuando esté indicado por motivos epidemiológicos. Es particularmente importante realizar un seguimiento continuo a los niños con diagnóstico temprano de FQ que permita detectar la colonización inicial por P. aeruginosa y la instauración de tratamientos antimicrobianos agresivos para prevenir la persistencia de la colonización inicial y retrasar la infección crónica17.
Además de la necesidad de utilizar medios selectivos específicos, otra recomendación particular es realizar siembras cuantitativas que permitan conocer la cantidad de los distintos microorganismos presentes en las secreciones respiratorias, un seguimiento más preciso de la evolución temporal del proceso de colonización crónica y valorar la eficacia de los distintos tratamientos instaurados33. Además, puede facilitar el aislamiento de patógenos que estén en baja proporción, así como la detección de un mayor número de morfotipos de P. aeruginosa. Desde el punto de vista metodológico, la siembra cuantitativa puede realizarse siguiendo los procedimientos convencionales de dilución logarítmica seriada de la muestra seguida de siembra por extensión en los medios indicados, aunque esta labor tediosa puede verse facilitada por el uso de sistemas semiautomáticos de siembra cuantitativa, como los sistemas de siembra en espiral.
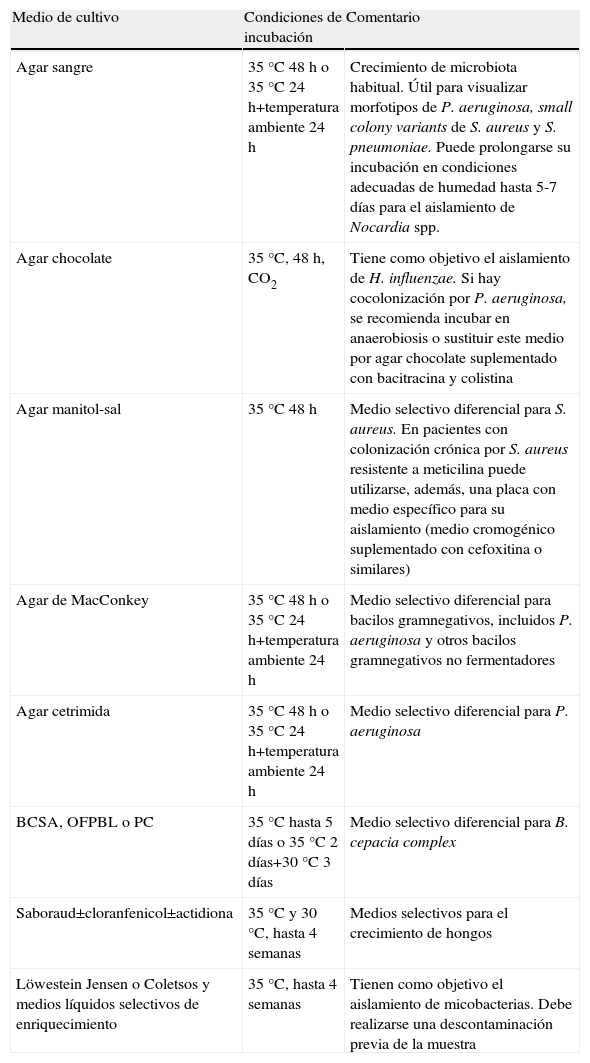
Medios de cultivo y condiciones de incubaciónEl cultivo microbiológico de las secreciones respiratorias de los pacientes con FQ debe incluir medios generales y selectivos-diferenciales para los patógenos habituales y una incubación algo más prolongada que la que se realiza con las muestras de otros pacientes. Con ello se facilita la recuperación de microorganismos que se encuentren en baja proporción, sobre todo cuando hay una colonización simultánea con P. aeruginosa. Se recomienda una incubación de al menos 48h, las primeras 24h a 35–37°C y luego a 30°C para facilitar el crecimiento de posibles bacilos gramnegativos no fermentadores. En algunos protocolos se recomienda específicamente la incubación de los medios entre 3 y 5 días. En la tabla 1 se indican los medios de cultivos más utilizados, las condiciones óptimas de incubación y su objetivo.
Medios de cultivo recomendables, condiciones óptimas de incubación y objetivos
| Medio de cultivo | Condiciones de incubación | Comentario |
| Agar sangre | 35°C 48h o 35°C 24h+temperatura ambiente 24 h | Crecimiento de microbiota habitual. Útil para visualizar morfotipos de P. aeruginosa, small colony variants de S. aureus y S. pneumoniae. Puede prolongarse su incubación en condiciones adecuadas de humedad hasta 5-7 días para el aislamiento de Nocardia spp. |
| Agar chocolate | 35°C, 48h, CO2 | Tiene como objetivo el aislamiento de H. influenzae. Si hay cocolonización por P. aeruginosa, se recomienda incubar en anaerobiosis o sustituir este medio por agar chocolate suplementado con bacitracina y colistina |
| Agar manitol-sal | 35°C 48 h | Medio selectivo diferencial para S. aureus. En pacientes con colonización crónica por S. aureus resistente a meticilina puede utilizarse, además, una placa con medio específico para su aislamiento (medio cromogénico suplementado con cefoxitina o similares) |
| Agar de MacConkey | 35°C 48h o 35°C 24h+temperatura ambiente 24 h | Medio selectivo diferencial para bacilos gramnegativos, incluidos P. aeruginosa y otros bacilos gramnegativos no fermentadores |
| Agar cetrimida | 35°C 48h o 35°C 24h+temperatura ambiente 24 h | Medio selectivo diferencial para P. aeruginosa |
| BCSA, OFPBL o PC | 35°C hasta 5 días o 35°C 2 días+30°C 3 días | Medio selectivo diferencial para B. cepacia complex |
| Saboraud±cloranfenicol±actidiona | 35°C y 30°C, hasta 4 semanas | Medios selectivos para el crecimiento de hongos |
| Löwestein Jensen o Coletsos y medios líquidos selectivos de enriquecimiento | 35°C, hasta 4 semanas | Tienen como objetivo el aislamiento de micobacterias. Debe realizarse una descontaminación previa de la muestra |
El cultivo de micobacterias requiere la utilización de protocolos y medios específicos y una comunicación expresa al laboratorio de microbiología de esta solicitud para establecer los cultivos y condiciones que aseguren su aislamiento. En todos los casos en que se solicite el cultivo para micobacterias es necesario realizar una baciloscopia mediante tinción de ácido-alcohol resistencia (Ziehl-Neelsen) o de fluorescencia adecuada para estos microorganismos (auramina-rodamina). Una tinción positiva no indica la presencia de M. tuberculosis ya que, como se indicó con anterioridad, este patógeno es infrecuente en el fibrótico quístico, no así las micobacterias atípicas. La frecuente presencia de P. aeruginosa en los esputos de estos pacientes requiere una descontaminación eficiente antes de su cultivo.
Los medios de cultivo empleados para los hongos no difieren de los utilizados habitualmente para muestras en las que se sospeche la presencia de estos microorganismos (tabla 1). Se ha recomendado la adición de gentamicina, amikacina o ciprofloxacino para inhibir la fuerte carga bacteriana que puede dificultar su crecimiento.
Consideraciones generales para la identificación de los agentes etiológicos en el contexto de la fibrosis quísticaEn términos generales, para identificar los agentes etiológicos implicados pueden seguirse los procedimientos empleados habitualmente en cualquier otro proceso infeccioso, aunque es importante tener en cuenta una serie de consideraciones específicas.
La identificación de S. aureus mediante las pruebas tradicionales de la coagulasa o la DNasa no presenta mayores problemas en el contexto de la FQ, aunque las morfologías coloniales atípicas (pequeño tamaño, no hemolíticas, no pigmentadas) de los mutantes SCV auxotróficos para timidina, típicos de esta enfermedad, pueden dificultar su reconocimiento en los medios de cultivo.
P. aeruginosa es, en principio, fácilmente reconocible en el laboratorio. No obstante, en los pacientes con FQ es muy frecuente encontrar múltiples morfologías coloniales atípicas entre las que cabe destacar las colonias mucoides o las SCV33. Otras morfologías coloniales frecuentes en la FQ son los morfotipos metálico y el rugoso. Además, es frecuente encontrar cepas no pigmentadas, carentes del pigmento verde característico; en otras ocasiones presentan otros diferentes, como ocurre en las cepas de pigmentación marrón, hiperproductoras de piomelanina33. Asimismo, la identificación basada en pruebas bioquímicas mediante galerías comerciales, como el API 20NE o las incluidas en los sistemas comerciales de microdilución para la identificación y el estudio de sensibilidad (MicroScan, Wider, Vitek, etc.), resulta a menudo dificultosa, principalmente debido al frecuentemente lento crecimiento en medio líquido de estas cepas, así como a su muchas veces disminuida capacidad de asimilación de los sustratos incluidos.
Teniendo en cuenta que B. cepacia es altamente transmisible, resistente a muchos antimicrobianos y puede conllevar un mal pronóstico, es importante usar métodos adecuados para su detección e identificación32. B. cepacia, especialmente cuando se obtiene de muestras respiratorias de pacientes con FQ, puede necesitar 3 días de incubación antes de que se vean las colonias en el medio selectivo. Las colonias en agar sangre o en los medios selectivos son lisas y ligeramente levantadas y, ocasionalmente, mucoides. Las colonias en MacConkey se ponen de rosa oscuro a rojo debido a la oxidación de lactosa en incubaciones prolongadas (4 a 7 días). La mayoría de los aislamientos son no pigmentados, pero en medios con hierro, como TSI, muchas cepas producen un pigmento amarillo brillante. Además, las colonias de B. cepacia tienen un olor característico. La mayoría de las cepas del complejo B. cepacia oxidan la sacarosa, el adonitol o ambos, mientras que R. pickettii no oxida ninguno de los dos. La mayoría de B. cepacia son lisina descarboxilasa positiva, mientras que no lo son R. pickettii, B. gladioli o Pandoraea spp. B. gladioli no oxida la lactosa, la maltosa y la sacarosa. Sin embargo, algunas B. cepacia de la variedad genómica III (B. cenocepacia) tampoco acidifican estos azúcares.
Los sistemas de identificación comerciales no son capaces de diferenciar entre las diferentes especies dentro del complejo y a menudo fallan en separar B. cepacia complex de otras especies o géneros relacionados como B. gladioli, R. pickettii, R. mannitolilytica y Pandoraea spp.33. En los estudios que han comparado diferentes sistemas comerciales, el API 20NE parece ser el mejor sistema comercial para identificar B. cepacia complex. Los sistemas automatizados como Vitek o MicroScan no son seguros para identificar estas bacterias. Se han usado diferentes métodos para identificar las variedades genómicas dentro del género Burkholderia. Las pruebas fenotípicas pueden ser capaces de diferenciar B. multivorans y B. stabilis. Entre las especies dentro del complejo hay algunas características que pueden ayudar a la identificación. Por ejemplo, B. multivorans, B. stabilis y B. dolosa no son capaces de oxidar la sacarosa. B. stabilis es ornitina descarboxilasa positiva, mientras que las otras dos son negativas. B. multivorans y B. dolosa se diferencian en la lisina descarboxilasa, pero es positiva en sólo el 53% de B. multivorans.
En cualquier caso, frecuentemente es difícil diferenciar estas especies mediante técnicas convencionales y se requiere la utilización de técnicas moleculares, descritas en apartados posteriores.
Estudio de la sensibilidad a los antibióticosPseudomonas aeruginosaLa periodicidad recomendada para la realización de estudios de sensibilidad a los antibióticos de las cepas de P. aeruginosa de pacientes con FQ es esencialmente la misma que la recomendada para la obtención de muestras respiratorias. En términos generales, la sensibilidad de los aislados procedentes de las exacerbaciones pulmonares suele coincidir con aquella obtenida en los últimos cultivos realizados en situación basal; por lo tanto, se puede utilizar estos datos para establecer las pautas empíricas iniciales. Debido a que las diferentes variantes fenotípicas de P. aeruginosa frecuentemente presentan distintos patrones de resistencia a los antibióticos, debe estudiarse la sensibilidad de todos los morfotipos detectados en los cultivos.
Respecto a las técnicas que se debe emplear, se consideran de referencia la dilución en agar y la microdilución, según las directrices del CLSI. Los sistemas comerciales de microdilución han logrado, en general, resultados poco satisfactorios, con altas tasas de errores muy graves (falsa sensibilidad), y por lo tanto no se recomienda utilizarlos34. Por el contrario, tanto la difusión con discos como el Etest (qué además permite determinar el valor preciso de CMI) han obtenido resultados satisfactorios en comparación con las técnicas de referencia, por esto la Fundación Americana de FQ recomienda enérgicamente su empleo como técnica sistemática en detrimento de los sistemas comerciales de microdilución. El Etest, además, permite determinar la CMI de tobramicina en un amplio intervalo de concentraciones y, por tanto, aplicar los dos puntos de corte propuestos para este antibiótico: el recomendado por el CLSI para administración por vía sistémica (resistente ⩾16μg/ml) y el recomendado recientemente para administración por vía inhalatoria (resistente ⩾128μg/ml)35. Independientemente de la técnica utilizada para el estudio de la sensibilidad, debe prolongarse la incubación de las placas al menos hasta 24h completas, y en los casos de variantes de lento crecimiento pueden ser necesarias hasta 48h de incubación. Es asimismo recomendable el uso de densidades bacterianas más elevadas para el estudio de las cepas mucosas (1 McFarland en lugar del 0,5 convencional), ya que el número de células viables, a igual densidad, es menor en estas cepas por la gran cantidad de alginato producido.
Finalmente, tanto la difusión con discos como el Etest, al contrario que la microdilución, permiten la detección de cepas hipermutadoras mediante la documentación de las subpoblaciones de mutantes resistentes (SMR) características de este tipo de cepas33,36. Según estudios previos, la presencia de SMR para al menos 3 de los antibióticos probados (ceftazidima, imipenem, meropenem, ciprofloxacino y tobramicina) puede utilizarse como criterio en la identificación de este tipo de cepas. La detección de cepas hipermutadoras puede ser de utilidad para el manejo clínico de la colonización-infección crónica por P. aeruginosa, ya que, como se ha comentado anteriormente, estas cepas son capaces de desarrollar resistencia rápidamente a la mayoría de los antibióticos y, por lo tanto, siempre se debe utilizar terapia combinada para su tratamiento.
Debido a la frecuente presencia de cepas de P. aeruginosa multirresistentes e incluso panresistentes en el contexto de la FQ, en ocasiones puede ser necesario recurrir al estudio de la actividad de combinaciones de antibióticos (estudios de sinergia) para la elección del tratamiento más adecuado. La técnica de referencia, aunque tediosa, es la microdilución en tablero de ajedrez. También se han evaluado técnicas basadas en el Etest, con resultados aceptables y, por lo tanto, aparentemente aplicables en la práctica habitual por su relativa sencillez. Otra técnica, utilizada actualmente en centros de referencia en Estados Unidos, determina la actividad bactericida de combinaciones de dos o más antibióticos (técnica denominada MCBT de las siglas inglesas multiple-combination bactericidal antibiotic testing). No obstante, su uso no mejoró la respuesta clínica en comparación con la elección de antimicrobianos basada en los estudios de sensibilidad convencionales, por lo que no se recomienda su utilización de forma sistemática, aunque puede resultar útil para casos particulares de infección por cepas multirresistentes o panresistentes17.
Otros microorganismosSin ánimo de pretender realizar una revisión exhaustiva de las técnicas, condiciones y connotaciones, más adecuadas para el estudio de la sensibilidad a los antibióticos de todos y cada uno de los patógenos potencialmente causantes de colonización-infección respiratoria crónica en los pacientes con FQ, este apartado se limitará a reseñar los aspectos diferenciales o de particular relevancia en el contexto de esta enfermedad.
Para el estudio de la sensibilidad a antibióticos de S. aureus de FQ pueden seguirse los procedimientos convencionales utilizados en otros tipos de infecciones. Cabe destacar, no obstante, la creciente relevancia del SARM en el contexto de FQ y, por lo tanto, la importancia de su correcta detección. Tanto el disco de cefoxitina, la siembra en medio de cribado con oxacilina como la aglutinación en látex para la detección de la PBP2a o la detección por PCR del gen mecA resultan adecuados para este propósito. La detección de la resistencia a meticilina por las técnicas basadas en el cultivo del microorganismo puede ser más compleja en los mutantes SCV, por lo tanto, es especialmente recomendable en estos casos la detección directa de la PBP2a o del gen mecA.
Respecto al estudio de la sensibilidad a antibióticos de H. influenzae, cabe destacar dos aspectos diferenciales característicos de las cepas obtenidas de pacientes con FQ. En primer lugar es importante tener en cuenta la relativamente alta prevalencia (>5%), al menos en nuestro medio, de cepas BLNAR. Por tanto, la evaluación de la resistencia a betalactámicos en este microorganismo no debe basarse únicamente en la detección de la producción de betalactamasa (estrategia recomendada en procedimientos de otros países); debe estudiarse la sensibilidad a ampicilina, preferiblemente por técnicas que permitan determinar la CMI, como la microdilución o el Etest. Otro aspecto relevante y característico de las cepas procedentes de pacientes con FQ es la alta prevalencia (que puede llegar hasta el 20%) de aislados resistentes (CMI>1μg/ml) a fluoroquinolonas, hecho extremadamente infrecuente fuera del contexto de esta enfermedad19. Es recomendable la determinación de la CMI (Etest o microdilución) para ciprofloxacino u otras fluoroquinolonas o la detección de la posible pérdida de sensibilidad a las fluroquinolonas con un disco de ácido nalidíxico.
Hasta la fecha no hay recomendaciones específicas en cuanto al método más adecuado para el estudio de la sensibilidad a antibióticos de B. cepacia complex, S. maltophilia, A. xylosoxidans u otros bacilos gramnegativos no fermentadores en el contexto de la FQ y, por lo tanto, debe seguirse la metodología habitualmente utilizada en cada laboratorio (difusión con discos, Etest o microdilución). Es importante considerar que estos microorganismos se caracterizan por presentar de forma natural resistencia a muchos de los antibióticos convencionalmente utilizados para el tratamiento de las infecciones por bacilos gramnegativos, y que esta resistencia es todavía mucho mayor en el contexto de la FQ. En la mayoría de los casos presentarán resistencia a todas las penicilinas, cefalosporinas, carbapenemes (todavía presentan cierta actividad frente a A. xylosoxidans), aminoglucósidos, ciprofloxacino e incluso a antibóticos de último recurso, como las polimixinas. Por el contrario, frecuentemente pueden ser sensibles a otros antibióticos de uso infrecuente en el tratamiento de las infecciones por bacilos gramnegativos y, generalmente, no evaluados de forma sistemática. Entre ellos, se encuentra el cotrimoxazol, la minociclina o las nuevas fluoroquinolonas; antibióticos que deben ser específicamente estudiados en caso de aislamiento de estos microorganismos. El cotrimoxazol, de hecho, se considera el tratamiento de elección en las infecciones por S. maltophilia, aunque la prevalencia de cepas resistentes procedentes de pacientes con FQ está aumentado notablemente. Por el contrario, todas las cepas estudiadas hasta la fecha son uniformemente sensibles a la minociclina. Las nuevas fluoroquinolonas, como el levofloxacino y el moxifloxacino, mejoran sustancialmente la actividad del ciprofloxacino, y pueden resultar una opción terapéutica de utilidad. Particularmente en el caso de B. cepacia, la limitación de las opciones terapéuticas frecuentemente hace necesario recurrir a la búsqueda de sinergias entre 2 o incluso 3 antimicrobianos; diversas combinaciones del meropenem, la tobramicina, el cotrimoxazol, el moxifloxacino o la rifampicina se han utilizado con este propósito.
Estudio de sensibilidad a los antibióticos en biofilmEl estudio estándar de sensibilidad a los antimicrobianos en el laboratorio de microbiología se realiza con un inóculo preparado a partir de microorganismos previamente crecidos hasta fase exponencial o a partir de colonias aisladas en placas de cultivo de 24–48h (NCCLS-M7A6). Este inóculo se denomina de crecimiento planktónico en contraposición al que se prepara a partir de bacterias desarrolladas en biopelículas o de crecimiento sésil. Este último se asemeja más al crecimiento que se produce en el pulmón del paciente con FQ. De hecho, diferentes autores han señalado que las discrepancias en la respuesta al tratamiento con los resultados que se obtienen en los estudios de sensibilidad podrían deberse a que éstos se realizan con un crecimiento inadecuado de los microorganismos.
Dentro de la complejidad que presenta el estudio de sensibilidad en biopelículas los protocolos que más se han utilizado son los que han derivado del dispositivo descrito por Ceri et al37, conocido como sistema de Calgary. El crecimiento de las bacterias se realiza en placas de microtitulación, idénticas a las que se utilizan para el estudio de sensibilidad por microdilución, cerradas con unas tapas que contienen púas (o pinchos) que se insertan en cada uno de los pocillos y sobre los que se desarrolla el biofilm. Una modificación de este sistema ha sido empleado por Moskowitz et al38 para el estudio de sensibilidad de P. aeruginosa procedentes de muestras respiratorias de pacientes con FQ y ha servido también como modelo para S. pneumoniae22. Esta modificación permite obtener un parámetro de actividad antimicrobiana, quizá más representativo para el tratamiento de la infección crónica que la propia CMI, denominado concentración inhibitoria de biopelículas (CIB).
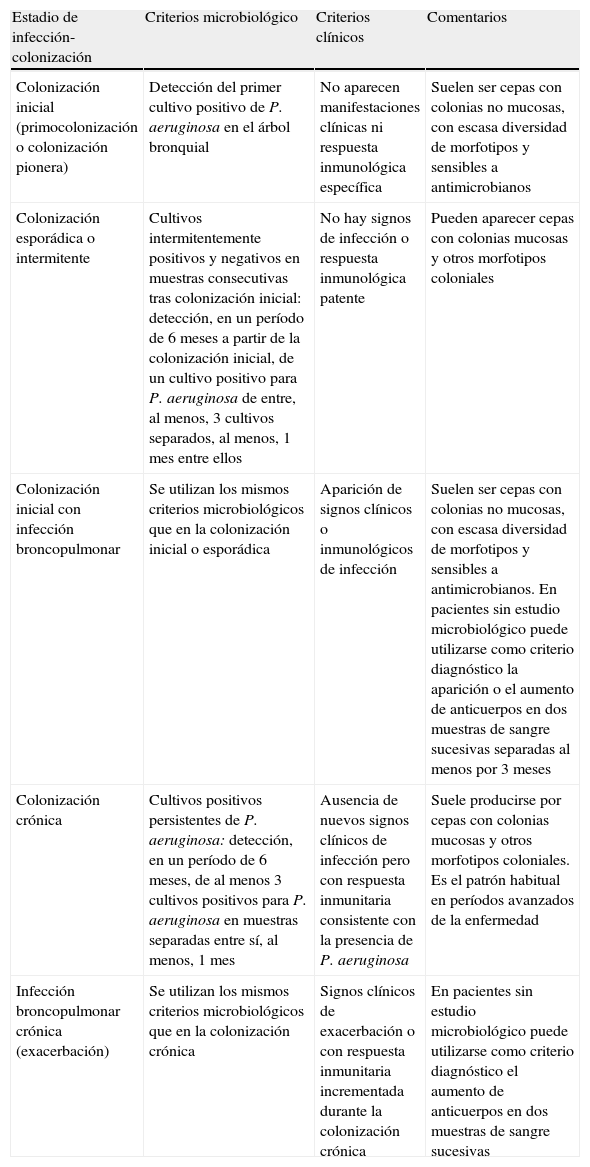
Interpretación e información de los cultivos cuantitativos secuencialesCasi todos los trabajos en los que se ha valorado la utilidad y las recomendaciones para la interpretación de los cultivos microbiológicos se han realizado para P. aeruginosa. La utilización de cultivos secuenciales de las secreciones respiratorias en los pacientes con FQ permite conocer el patrón de colonización y definir el estadio de colonización broncopulmonar en el que se encuentra el paciente (primocolonización, colonización intermitente, colonización crónica) y confirmar, desde el punto de vista microbiológico, la presencia de exacerbaciones. Asimismo, la realización de recuentos bacterianos en el cultivo facilita la medición de la respuesta al tratamiento en los pacientes con colonización crónica y documenta las exacerbaciones. Aunque hay controversias al respecto, el incremento de los recuentos bacterianos y el aumento del número de morfotipos de P. aeruginosa se han relacionado con un deterioro de la función pulmonar. También se ha evidenciado en los pacientes con colonización por S. maltophilia. En la tabla 2 se recogen, de manera sintética, las definiciones realizadas por el Grupo Español de Consenso del Tratamiento Antimicrobiano en el Paciente con Fibrosis Quística y los criterios microbiológicos establecidos en la colonización-infección pulmonar por P. aeruginosa39. Su objetivo es la clasificación de los pacientes para establecer pautas de tratamiento antimicrobiano. Por el momento no hay criterios publicados que definan los estadios de colonización por otros patógenos.
Estadios de infección-colonización por P. aeruginosa y criterios microbiológicos de valoración
| Estadio de infección-colonización | Criterios microbiológico | Criterios clínicos | Comentarios |
| Colonización inicial (primocolonización o colonización pionera) | Detección del primer cultivo positivo de P. aeruginosa en el árbol bronquial | No aparecen manifestaciones clínicas ni respuesta inmunológica específica | Suelen ser cepas con colonias no mucosas, con escasa diversidad de morfotipos y sensibles a antimicrobianos |
| Colonización esporádica o intermitente | Cultivos intermitentemente positivos y negativos en muestras consecutivas tras colonización inicial: detección, en un período de 6 meses a partir de la colonización inicial, de un cultivo positivo para P. aeruginosa de entre, al menos, 3 cultivos separados, al menos, 1 mes entre ellos | No hay signos de infección o respuesta inmunológica patente | Pueden aparecer cepas con colonias mucosas y otros morfotipos coloniales |
| Colonización inicial con infección broncopulmonar | Se utilizan los mismos criterios microbiológicos que en la colonización inicial o esporádica | Aparición de signos clínicos o inmunológicos de infección | Suelen ser cepas con colonias no mucosas, con escasa diversidad de morfotipos y sensibles a antimicrobianos. En pacientes sin estudio microbiológico puede utilizarse como criterio diagnóstico la aparición o el aumento de anticuerpos en dos muestras de sangre sucesivas separadas al menos por 3 meses |
| Colonización crónica | Cultivos positivos persistentes de P. aeruginosa: detección, en un período de 6 meses, de al menos 3 cultivos positivos para P. aeruginosa en muestras separadas entre sí, al menos, 1 mes | Ausencia de nuevos signos clínicos de infección pero con respuesta inmunitaria consistente con la presencia de P. aeruginosa | Suele producirse por cepas con colonias mucosas y otros morfotipos coloniales. Es el patrón habitual en períodos avanzados de la enfermedad |
| Infección broncopulmonar crónica (exacerbación) | Se utilizan los mismos criterios microbiológicos que en la colonización crónica | Signos clínicos de exacerbación o con respuesta inmunitaria incrementada durante la colonización crónica | En pacientes sin estudio microbiológico puede utilizarse como criterio diagnóstico el aumento de anticuerpos en dos muestras de sangre sucesivas |
La identificación correcta de B. cepacia probablemente es uno de los aspectos más relevantes en el control microbiológico de los pacientes con FQ. El fallo en el reconocimiento de este importante patógeno puede tener notables consecuencias en la evolución clínica del paciente, mientras que la falsa identificación de otros microorganismos, como B. cepacia, conlleva también un elevado impacto médico, social y psicológico. Asimismo, la importancia clínica de las diferentes variedades genómicas no es la misma por la que se debería realizar la identificación de la variedad dentro del complejo B. cepacia. Cuando se aísla este patógeno, debe informarse lo más rápidamente posible, y confirmar el resultado en un centro de referencia. Asimismo, se deben aplicar medidas de control estrictas, particularmente es necesario separar a estos pacientes de otros no colonizados para evitar la transmisión cruzada. Se recomienda también mantener separados entre sí a los pacientes colonizados por B. cepacia, para evitar la transmisión de cepas particularmente virulentas.
El informe del laboratorio de microbiología para las muestras respiratorias de pacientes con FQ debe incluir la identificación y el recuento (logaritmo de ufc/ml de esputo) de todos los microorganismos potencialmente patógenos presentes, y su antibiograma correspondiente. En el caso de P. aeruginosa, se debe especificar el número de morfotipos distintos presentes y se debe incluir un antibiograma para cada uno de ellos. Particularmente, debe especificarse, en su caso, la presencia de cepas con morfotipo mucoso. En el informe del antibiograma se debe aplicar, como en cualquier otro tipo de infección, los puntos de corte establecidos para la administración de los antibióticos por vía sistémica; también es recomendable aplicar puntos de corte específicos para la vía inhalada. Actualmente, sólo hay recomendaciones específicas para la tobramicina, que es el antibiótico más frecuentemente usado como terapia de mantenimiento por vía inhalatoria en los pacientes con FQ35. Los puntos de corte establecidos indican considerar como potencialmente tratables por esta vía las cepas cuya CMI de tobramicina sea ⩽64μg/ml (puntos de corte convencionales del CLSI ⩽4μg/ml sensible).
En función de la experiencia de cada centro, los estudios de sensibilidad convencionales pueden completarse con la determinación de otros parámetros descritos en apartados anteriores: determinación de la concentración inhibitoria de biopelículas (CIB) y la concentración inhibitoria de mutantes (CPM). Si bien estos parámetros no pueden sustituir, hoy por hoy, a la CMI clásica, pueden aportar información complementaria útil en la selección de los regímenes antibióticos más adecuados para el tratamiento de la infección crónica por P. aeruginosa. De igual forma, sería recomendable realizar e informar los estudios de sinergia en las cepas multirresistentes.
Asimismo, en función de la experiencia de cada centro, debe valorarse la detección de cepas hipermutadoras y, en su caso, informar de su aislamiento y añadir en las observaciones del informe una llamada a la necesidad de utilizar combinaciones de antibióticos para su tratamiento debido al elevado riesgo de desarrollar resistencia. Una de las cuestiones más relevantes que surgen al examinar los antibiogramas de las cepas hipermutadoras es cómo interpretar e informar los resultados de sensibilidad por la frecuente presencia de SMR. La aproximación más conservadora sería considerar las subpoblaciones resistentes y leer las CMI (Etest) o diámetros de los halos de inhibición (difusión con discos) que producen una completa inhibición del crecimiento. El problema de esta consideración es que en muchos casos informaríamos la cepa como resistente a todos los antibióticos (incluso aunque no presente mecanismos de resistencia adquiridos) y, por lo tanto, no habría ninguna opción terapéutica disponible. Sin embargo, los estudios de sinergia demuestran que las SMR desaparecen cuando se utilizan combinaciones de antibióticos con distinto mecanismo de resistencia. Resulta importante distinguir la verdadera resistencia de la debida a la presencia de SMR producida por la propia frecuencia de mutación incrementada, ya que al contrario de lo que ocurre con el primer tipo de resistencia, el segundo podría ser potencialmente combatido con combinaciones de antibióticos. En este sentido, recientemente se ha propuesto una alternativa para la interpretación de los antibiogramas (difusión con discos y Etest) de las cepas hipermutadoras33,36: a) leer las CMI (o diámetros de los halos de inhibición) de la población general y aplicar los puntos de corte definidos para las categorías de sensibilidad (S, I, R), y b) examinar la presencia de SMR y leer las CMI o diámetros de los halos de inhibición producidos por el crecimiento de éstas y aplicar los puntos de corte definidos para las categorías de sensibilidad (S, I, R). La lectura definitiva de las SMR puede requerir 12–24h adicionales de incubación en las cepas de lento crecimiento. Informar de los antibióticos para los cuales la población general es sensible, pero que contiene SMR que supera los puntos de corte como «resistente, pero potencialmente activo para su uso en terapia combinada».
Técnicas rápidas de diagnósticoDetección de P. aeruginosa y otros patógenos en muestras respiratorias por métodos molecularesEl cultivo es la técnica utilizada habitualmente para detección de P. aeruginosa y otros bacilos gramnegativos. Sin embargo, en ocasiones el cultivo es negativo (especialmente en pacientes con tratamiento antimicrobiano o con baja carga bacteriana) o se requieren varios días para llegar a la identificación. Por eso, cada vez es más frecuente la utilización de técnicas moleculares que permiten un diagnóstico más rápido y con menor límite de detección40.
La amplificación mediante PCR de los genes ribosomales, la posterior secuenciación y comparación con bases de datos, permite identificar prácticamente todos los microorganismos y ha permitido identificar nuevas especies. En los últimos años se ha utilizado con éxito la técnica de FISH (fluorescent in situ hybridization) para detección de los principales patógenos en pacientes con FQ (P. aeruginosa, S. aureus, H. influenzae, B. cepacia, Burkholderia spp. y S. maltophilia). La técnica permite obtener resultados en pocas horas y se ha descrito muy buena especificidad, pero sensibilidad del 90%, comparada con el cultivo, ya que requiere alta densidad de colonización para dar un resultado positivo.
Las técnicas de PCR utilizadas para detectar P. aeruginosa en esputo de pacientes con FQ presentan, en general, alta sensibilidad (del 93 al 100% según los primers que se utilicen)40. Entre los genes utilizados como diana encontramos el ARNr 16S, oprI y oprL, algD o el gen de la exotoxina A (ETA). Uno de los genes más utilizados, oprI, está muy conservado entre los diferentes miembros de Pseudomonas, y el gen oprL es específico de P. aeruginosa, incluidas las colonias no pigmentadas y las mucoides. Por lo tanto, es posible realizar una PCR múltiple que detectará P. aeruginosa y otras Pseudomonas en la misma reacción. La técnica aplicada a muestras clínicas muestra una alta sensibilidad, ya que es capaz de detectar 100ufc/ml. Estas técnicas permiten, además, detectar colonización con P. aeruginosa una media de 4,5 meses antes que el cultivo.
Las técnicas moleculares también se han utilizado con éxito para detectar B. cepacia complex a partir de muestras respiratorias de pacientes con FQ, mediante diferentes primers diseñados para amplificar el gen recA, regiones conservadas del gen 16S ARNr, o la región espaciadora 16S a 23S41,42. Los primeros estudios se realizaron con los primers para el 16S ARNr; sin embargo, estos primers estaban diseñados frente a una cepa de B. cepacia de la variedad genómica I y actualmente se sabe que tienen baja especificidad para B. multivorans y B. vietnamiensis y, además, pueden tener reacción cruzada con otras bacterias distintas de B. cepacia complex. Los primers de la región espaciadora 16S-23S permiten identificar sólo 3 variedades genómicas (B. cepacia, B. cenocepacia y B. stabilis) por lo que no son útiles para detectar todas las cepas del complejo. Los primers del gen recA permiten detectar bacterias del complejo B. cepacia con alta especificidad y posteriormente identifican las diferentes variedades genómicas por RFLP41. De hecho, los casos descritos de PCR negativa y cultivo positivo para B. cepacia resultaron ser cepas de Burkholderia pero no del complejo cepacia. El método es poco sensible, pues necesita 106ufc/g de esputo para dar un resultado positivo, debido a que se amplifica un fragmento relativamente grande (para poder obtener patrones diferentes de digestión) y en el genoma de B. cepacia hay una única copia del gen recA. El límite de detección es más bajo cuando se usan los primers de la región espaciadora del 16S-23S, debido a que en el genoma hay múltiples copias del operón ARNr. Sin embargo, a pesar de que el límite de detección por gramo de esputo es alto, los resultados clínicos son excelentes y muestra una muy buena correlación con los datos obtenidos por cultivo.
Técnicas para el estudio directo de la sensibilidad y la resistencia a los antibióticosEn los últimos años cada vez cobra más interés el desarrollo de procedimientos que permitan conocer la sensibilidad a antibióticos directamente en la siembra primaria. La principal motivación para esta aproximación reside en que con los procedimientos convencionales hay un retraso importante en la obtención de los resultados, no menos de 2 ó 3 días después de la toma de muestra en el mejor de los casos, lo que limita enormemente el impacto positivo de la terapia dirigida, particularmente en las infecciones graves de rápida evolución. Según estudios recientes, la realización de un antibiograma por Etest directamente de las muestras respiratorias de los pacientes con neumonía relacionada con ventilación mecánica ofrece resultados en 18–24h prácticamente concordantes en su totalidad con el antibiograma convencional. En primera instancia cabría suponer que la disminución del tiempo necesario para la obtención de los resultados no debe ser aparentemente una prioridad imperiosa en el contexto de la colonización-infección crónica en pacientes con FQ, y que por tanto no estaría justificada esta aproximación. No obstante, existe otra connotación que quizá abogue a favor del estudio directo de sensibilidad: la gran heterogeneidad poblacional en las infecciones crónicas. Esta marcada heterogeneidad obliga al estudio individual de cada uno de los morfotipos presentes en las muestras (ya que frecuentemente presentan diferente sensibilidad), con el aumento notable del coste y la complejidad del procedimiento; es más, ello no garantiza la detección de poblaciones resistentes, ya que colonias pertenecientes al mismo morfotipo también podrían presentar distinto perfil de resistencia a antimicrobianos. Principalmente son dos las técnicas propuestas para este fin, el Etest directo y la siembra en medios con antibióticos43. Si bien sus potenciales prestaciones son prometedoras, no están suficientemente validadas por estudios comparativos y por eso ninguna de ellas puede sustituir actualmente al procedimiento convencional, aunque pueden ofrecer resultados complementarios de utilidad.
Procedimientos adicionales a realizar en situaciones especialesTécnicas moleculares para la identificación de bacilos gramnegativos no fermentadoresComo se ha comentado anteriormente, la identificación de B. cepacia y otros bacilos gramnegativos no fermentadores requiere de técnicas moleculares, ya que los métodos fenotípicos no son suficientemente fiables y se puede realizar identificación incorrecta de bacterias con alta trascendencia clínica, como P. aeruginosa o, sobre todo, B. cepacia.
La técnica molecular definitiva es la secuenciación, que permite identificar cualquier bacteria comparando la secuencia en la base de datos o incluso permite identificar bacterias nuevas. La comparación de la secuencia del gen ARNr 16S se ha usado como prueba de referencia en estudios filogenéticos de bacterias; sin embargo, la resolución puede ser demasiado baja para distinguir especies muy relacionadas. También se ha utilizado la secuencia del gen gyrB como complementaria a la del ARNr para identificar algunas bacterias como Pandoraea spp.
Si la secuenciación no está disponible, se puede realizar el patrón de restricción del fragmento amplificado (PCR-RFLP) o el polimorfismo de la conformación de la cadena simple (SSCP), que puede ser analizado mediante electroforesis en gel de poliacrilamida no desnaturalizante o electroforesis capilar. La técnica de ARDRA (amplified ribosomal DNA restriction analysis), en la que se amplifica el gen y posteriormente se realiza una digestión con enzimas de restricción, ha demostrado ser una alternativa a la secuenciación para diferenciar, por ejemplo, Inquilinus spp. o Pandoraea spp. de otros bacilos gramnegativos no fermentadores.
La técnica de AFLP (amplified fragment length polymorphism) convencional puede ser útil para identificar B. cepacia, pero es técnicamente compleja. La PCR en tiempo real está accesible en muchos laboratorios clínicos, pero no es útil para detectar cualquier microorganismo presente, sino que se utiliza para identificar un género o especie según la amplificación de un gen específico42. Se han estudiado diferentes dianas y la mejor identificación, especialmente en la identificación de P. aeruginosa atípicas, se consigue cuando se detectan varios genes simultáneamente.
La determinación de las variedades genómicas de B. cepacia se basa en un estudio taxonómico complejo que incluye pruebas fenotípicas, como el perfil de proteínas totales, y genotípicas, como hibridación ADN-ADN. Recientemente, se están utilizando con éxito las pruebas basadas en PCR para identificar las variedades genómicas, principalmente basadas en el gen ARNr 16S o en el recA41,42. La amplificación del gen ARNr 16S, y posterior análisis mediante secuenciación o con enzimas de restricción, tiene un uso limitado para diferenciar las variedades genómicas, ya que no presenta suficiente variabilidad. El estudio del gen recA se puede afrontar desde diferentes estrategias que dependerán de la disposición de medios en cada centro: a) amplificación del gen con primers específicos que permite identificar un aislamiento como perteneciente al complejo B. cepacia; b) digestión del fragmento amplificado con HaeIII y con MnlI que permite clasificarlos dentro de las principales variedades; c) PCR específica de cada variedad de forma individualizada, y d) secuenciación del fragmento amplificado, que permite la identificación y la clasificación precisa.
Técnicas de epidemiología molecular para el seguimiento de la colonización-infección broncopulmonar crónicaLa aplicación de las técnicas de tipificación molecular puede contribuir de manera notable a la optimización del seguimiento microbiológico de la infección-colonización crónica por P. aeruginosa en pacientes con FQ. La electroforesis en campo pulsado (ECP), como para la mayoría de los microorganismos, es la técnica de referencia, aunque aquellas basadas en la PCR (REP, AP, ERIC o RAPD, entre otras) pueden resultar, por su rapidez y sencillez, más fácilmente adaptables al trabajo sistemático del laboratorio de microbiología.
Una de las principales aportaciones de las técnicas de tipificación molecular es su utilidad para el propio diagnostico de la infección-colonización crónica. Como se ha comentado en apartados anteriores, los primeros estadios del proceso suelen cursar con aislamiento esporádico o intermitente de P. aeruginosa; la tipificación molecular puede permitir determinar si realmente se trata ya de una colonización crónica, todavía con baja carga bacteriana, o si por el contrario se trata únicamente de colonizaciones esporádicas por cepas ambientales diferentes. De igual forma, las técnicas de tipificación molecular pueden resultar de gran utilidad para la evaluación de la eficacia de los tratamientos erradicadores iniciales. Si bien es factible conseguir en estos primeros estadios la negativización de los cultivos por un periodo más o menos prolongado, es frecuente que al cabo de los meses se vuelva a documentar cultivos positivos: la tipificación molecular nos permitirá determinar si nos enfrentamos a una falsa erradicación (la cepa original persistió en densidades por debajo del umbral de detección) o si, por el contrario, nos encontramos ante una nueva colonización-infección por otra cepa diferente (y por lo tanto el tratamiento erradicador fue efectivo)39.
En los últimos años se ha documentado la diseminación epidémica de cepas concretas de P. aeruginosa entre los pacientes con FQ7. El primer clon epidémico fue originalmente descrito en 1995 en Liverpool (LES, Liverpool epidemic strain) y actualmente se encuentra diseminado por todo el Reino Unido. De igual forma, se ha descrito una notable transmisión de clones epidémicos entre pacientes con FQ en Australia. Finalmente, en un estudio reciente se documenta la transmisión cruzada entre pacientes con FQ de dos clones concretos mayoritarios durante más de dos décadas en Dinamarca; estos clones aparentemente estarían adaptados no sólo para producir infección crónica, sino también para transmitirse de forma eficiente entre los pacientes con FQ. Frecuentemente, además, estos clones epidémicos son resistentes a múltiples antibióticos y su aislamiento conlleva un peor pronóstico de la infección crónica. Por lo tanto, es recomendable mantener una vigilancia activa para la detección y la contención de clones potencialmente epidémicos.
Diagnóstico serológico en el contexto de la FQLa diferenciación entre colonización, infección y enfermedad no es fácil de establecer en el caso de algunos de los microorganismos oportunistas que afectan de forma crónica a los pacientes con FQ. En determinados casos es importante detectar cuanto antes la infección inicial ya que el tratamiento antibiótico precoz es útil para evitar el establecimiento de la infección crónica antes de que sea imposible erradicarla. En otros casos, la distinción entre colonización e infección, aunque difícil, puede establecerse según criterios basados en la frecuencia o la persistencia de su aislamiento. El diagnóstico de enfermedad activa o deterioro pulmonar atribuible a ellos es muy difícil en estos pacientes basado en criterios clínicos, funcionales o radiológicos porque prácticamente todos los pacientes con FQ presentan alteraciones debidas a la propia enfermedad. En este contexto, el estudio de la respuesta de anticuerpos específicos contra microorganismos, como Aspergillus, P aeruginosa o MNT, es un complemento útil para la detección precoz de la colonización, la diferenciación entre infección y enfermedad activa, como marcador pronóstico, para la toma de decisiones terapéuticas o, en algún caso, para el seguimiento de la respuesta al tratamiento33. En el caso de Aspergillus, la presencia de anticuerpos específicos es un criterio fundamental, aunque no suficiente, para confirmar o excluir el diagnóstico de ABPA30. Estudios longitudinales de pacientes con FQ desde el nacimiento han mostrado que la detección de anticuerpos específicos anti-P. aeruginosa puede preceder entre 3 y 24 meses al primer aislamiento por cultivo. Una vez aislada por primera vez, los títulos elevados o su aumento pueden preceder hasta en 3 años a la instauración de la infección crónica44. La detección de anticuerpos contra P. aeruginosa sería útil para el diagnóstico de la infección inicial por este microorganismo, sobre todo cuando no es posible la obtención de muestras adecuadas para cultivo, principalmente en niños muy pequeños45.

