




Con la finalidad de identificar afinidades biológicas entre poblaciones del sitio arqueológico de Teotihuacán, se efectuó un análisis morfológico de los senos frontales de un total de 125 individuos adultos prehispánicos de ambos sexos exhumados en diferentes exploraciones. La muestra incluye un sector de los sujetos sacrificados en el Templo de la Serpiente Emplumada de Teotihuacan, otro del Posclásico, y cráneos mayas hallados en la Cueva de las Banquetas. Asimismo se usó una muestra control de 14 habitantes de la Ciudad de México mediante el uso de tomografía.
Los datos morfológicos fueron analizados usando la técnica de morfometría geométrica. Se observó que la población del Posclásico se separa completamente de las poblaciones Clásicas de Teotihuacan (Estado de México) y la Cueva de las Banquetas (Chiapas), en tanto que los individuos encontrados en el Templo de la Serpiente Emplumada presentan una mayor afinidad con las poblaciones mayas que con la población teotihuacana. En lo referente a la variación que presenta cada grupo, se observó que la muestra de Teotihuacan es la que presenta más dispersión en la morfología de los senos frontales. Este dato concuerda con el papel que desempeñó esta urbe como centro económico, ideológico y cultural. La investigación muestra que el estudio de los senos frontales es un marcador fiable para estudiar afinidades biológicas en poblaciones antiguas. Una de sus ventajas es que es factible incluir tanto cráneos completos como fragmentos de huesos frontales, ampliando así el tamaño de la muestra.
The morphological analysis of the frontal sinuses of 125 pre-Columbian human adults of both sexes exhumed in different explorations allowed to identify biological affinities between populations at the archaeological site of Teotihuacan. The sample included subjects sacrificed in the Temple of the Feathered Serpent at Teotihuacan and Postclassic subjects, as well as Mayan skulls found in the Cueva de las Banquetas. A control sample of 14 inhabitants of Mexico City was also used, assessed by tomography.
The morphological data were analysed using geometric morphometric methods. It was observed that the Postclassic population had absolutely no affinity with the Classic populations of Teotihuacan (State of Mexico) and the Cueva de las Banquetas (Chiapas), while the subjects found in the Temple of the Feathered Serpent had greater affinity with the Mayan populations than with the Teotihuacan population. Regarding the variation of each group, the Teotihuacan sample was found to have the greatest dispersion in terms of the morphology of the frontal sinuses. These data are consistent with the role played by this city as an economic, ideological and cultural centre. This research shows that the study of the frontal sinuses is a reliable marker in to establish biological affinities among ancient populations. One of its advantages is that it is feasible to include both complete skulls and fragments of frontal bones, thus enlarging the sample size.
La Ciudad de Teotihuacan ha sido reconocida como un importante núcleo que marcó pautas en la organización política, económica y religiosa durante el horizonte Clásico de la cuenca México. Además, era un punto de intercambio entre el Norte, el Centro de México y Centroamérica, y es innegable que debió estar conformada por una composición multiétnica (Arnauld, Manzanilla y Smith, 2012; Álvarez-Sandoval, Manzanilla, González-Ruiz, Malgosa y Montiel, 2015; Manzanilla, 2015). No obstante, ha sido difícil abordar el tema de la conformación biológica de los teotihuacanos debido a que los restos óseos humanos recuperados durante las excavaciones en su mayoría se localizan en malas condiciones de preservación. Esto en gran medida es resultado de las prácticas funerarias y el manejo de los cadáveres que ha generado conservación diferencial. El ejemplo más claro son las segundas exequias, la reutilización de los espacios funerarios y el uso de los huesos como materia prima. Tales aspectos imposibilitan el estudio métrico de los materiales por la fragmentación de los elementos, aunado al problema de la diagénesis de los huesos que dificulta la posibilidad de obtener información molecular a partir de ellos. No se puede solucionar el problema de la conservación de materiales, sin embargo, se puede solventar la cuestión de cómo estudiar materiales tan destruidos. Para esto, se propone utilizar un rasgo anatómico que permite realizar estudios morfológicos comparativos para establecer afinidad biológica entre restos óseos humanos hallados en Teotihuacan. Debido a que resulta ser un elemento fiable como marcador de parentesco biológico, se eligió a los senos frontales (SF) como rasgo diagnóstico (Kondrat, 1995; Kirk, Wood y Goldstein, 2002; Cameriere, Ferrante, Molleson y Brown, 2008). Su registro y posterior análisis se puede realizar en cráneos completos, incompletos y fragmentados debido a que el punto básico de representación es el hueso frontal (Meza, 2006; Meza Peñaloza, 2015).
Se puede afirmar que los SF han sido de gran utilidad en investigaciones forenses debido a que este rasgo es distintivo en cada persona en cuanto a forma y tamaño. Coloquialmente se les conoce como las huellas dactilares del cráneo (Johnson y Kennedy, 1961; Blanton y Biggs, 1969; Szilvassy, 1973; Ubelaker, 1984; Krogman e Iscan, 1986; Yoshino, Miyasaka, Sato y Seta, 1987; Hauser y de Stefano, 1989; Reichs, 1993; Donald, 1995; Quatrehomme et al., 1996; Bergman, Thompson, Afifi y Saadeh, 1997; Nambiar, Naidu y Subramaniam, 1999; Kirk et al., 2002). Asimismo se ha considerado a este rasgo como un excelente marcador biológico que se puede aplicar en el estudio de colecciones osteológicas de poblaciones antiguas y modernas. Nótese que los trabajos realizados en cráneos procedentes de la edad de bronce de la época romana y medieval en Inglaterra y entre grupos de esquimales modernos están en concordancia (Buckland-Wright, 1970; Greene y Armelagos, 1972; Greene y Scott, 1973; Hanson y Owsley, 1980; Kondrat, 1995; Vinyard y Smith, 1997; Koppe y Nagai, 1999; Lynnerup, Homøe y Skovgaard, 1999). Análisis previos realizados en muestras provenientes de contextos domésticos y ceremoniales de Teotihuacan demostraron la fiabilidad del uso de SF como rasgo diagnóstico para establecer parámetros de afinidad biológica entre sectores de la población inhumados bajo los pisos de las unidades habitacionales, hallados en basureros o en contextos sacrificiales (Meza Peñaloza, 2015). En este trabajo estudiaremos los patrones de variación en el contorno de SF para establecer la disparidad al interior de cada una de las poblaciones y estimar distancias biológicas entre las poblaciones prehispánicas. Se seleccionaron muestras del Clásico mesoamericano, teotihuacanos, mayas y sacrificados del templo de la Serpiente Emplumada; una muestra del Posclásico asentada en Teotihuacan tras el colapso de esta urbe, y una muestra contemporánea de la Ciudad de México con edades y sexos conocidos que no presentaban lesiones o enfermedades que pudieran afectar la forma y el crecimiento de los SF.
Métodos y técnicasLa muestraCorresponde a individuos adultos de ambos sexos. Para estimar edad y sexo en los individuos prehispánicos se aplicaron técnicas morfológicas estandarizadas (Buikstra y Ubelaker, 1994; White, Black y Folkens, 2012). Para la población moderna se cuenta con la información directa de cada uno de los voluntarios.
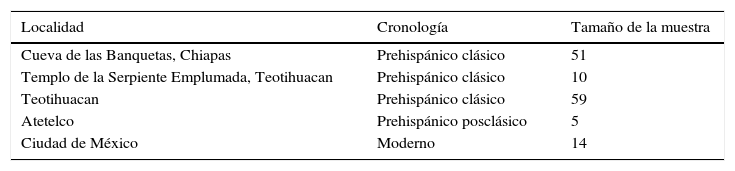
La población prehispánica está representada por 125 sujetos: 59 provenientes en su mayoría de contextos domésticos de Teotihuacan que corresponden al período de mayor desarrollo de la ciudad (200-600d.C.), 10 representantes de los sacrificados del Templo de la Serpiente Emplumada, 5 fueron hallados en contextos habitacionales datados para el periodo Posclásico (1000-1100d.C.) (Kanjou y Andrade, 2003). Los 51 cráneos de origen maya fueron hallados en la Cueva de las Banquetas, en Chiapas, que fue utilizada como recinto funerario durante el periodo Clásico (Romano Pacheco, Martínez Bautista y Jaén Esquivel, 2011; Tiesler, 2014). Adicionalmente se contó con datos de población actual de habitantes de la Ciudad de México representada por 14 sujetos (cuadro 1).
Distribución de la muestra de acuerdo a región y cronología
| Localidad | Cronología | Tamaño de la muestra |
|---|---|---|
| Cueva de las Banquetas, Chiapas | Prehispánico clásico | 51 |
| Templo de la Serpiente Emplumada, Teotihuacan | Prehispánico clásico | 10 |
| Teotihuacan | Prehispánico clásico | 59 |
| Atetelco | Prehispánico posclásico | 5 |
| Ciudad de México | Moderno | 14 |
Este grupo está integrado por 59 individuos hallados en contextos habitacionales explorados dentro de los proyectos de la VentillaB, Ventilla 92-9, Tetitla, Libramiento Pirámides y Mazapa y por entierros asociados a la Plaza de la Luna, Calzada de los Muertos y otras excavaciones de salvamento realizados por personal del INAH durante la década de 1960 y las exploraciones concernientes al Proyecto Arqueológico Teotihuacan 1980-1982 (para más información, revisar Meza Peñaloza, 2015).
Templo de la Serpiente EmplumadaSe estudiaron los entierros 16 (individuos A, B y C), 204 (individuos C, E, F, G, I, J, K, LL, N) y la Sepultura NW1, integrada por un único individuo. Todos ellos se encontraron al norte de la estructura. Sugiyama (2005) los identifica como entierros periféricos contemporáneos al final de la edificación de la pirámide.
Atetelco PosclásicoEl conjunto Atetelco se localiza a 1km al este de la Calzada de los Muertos. Consta de espacios habitacionales y lugares dedicados a labores institucionales, correspondientes a las fases Coyotlatelco, Mazapa y Azteca (Kanjou y Andrade, 2003).
Población contemporáneaEstá representada por mestizos de la Ciudad de México. Se utilizaron las tomografías de 14 personas mayores a 15años para poder comparar la forma y el tamaño de los SF entre ambos sexos (7 masculinos, 7 femeninos). En ningún caso los participantes presentaban cuadros de infección, traumatismos o características congénitas que pudieran afectar la forma o el tamaño de los SF y en todos los casos se contó con el consentimiento informado de cada uno de los participantes para acceder a la información relacionada con ancestría y el uso que se daría a las imágenes obtenidas de las tomografías.
Cueva de las BanquetasTras la denuncia de saqueo en 1983 se llevó a cabo el rescate arqueológico de la Cueva de las Banquetas, localizada en el municipio de la Trinitaria, Chiapas (coordenadas 16°05′ latitud Norte y 91°58′ longitud Oeste). De acuerdo a los reportes preliminares, la cerámica asociada corresponde al Clásico tardío o quizás al Posclásico temprano (800-1100d.C.). Análisis del modelado cefálico identifican a esta muestra dentro del periodo Clásico (Tiesler, 2014).
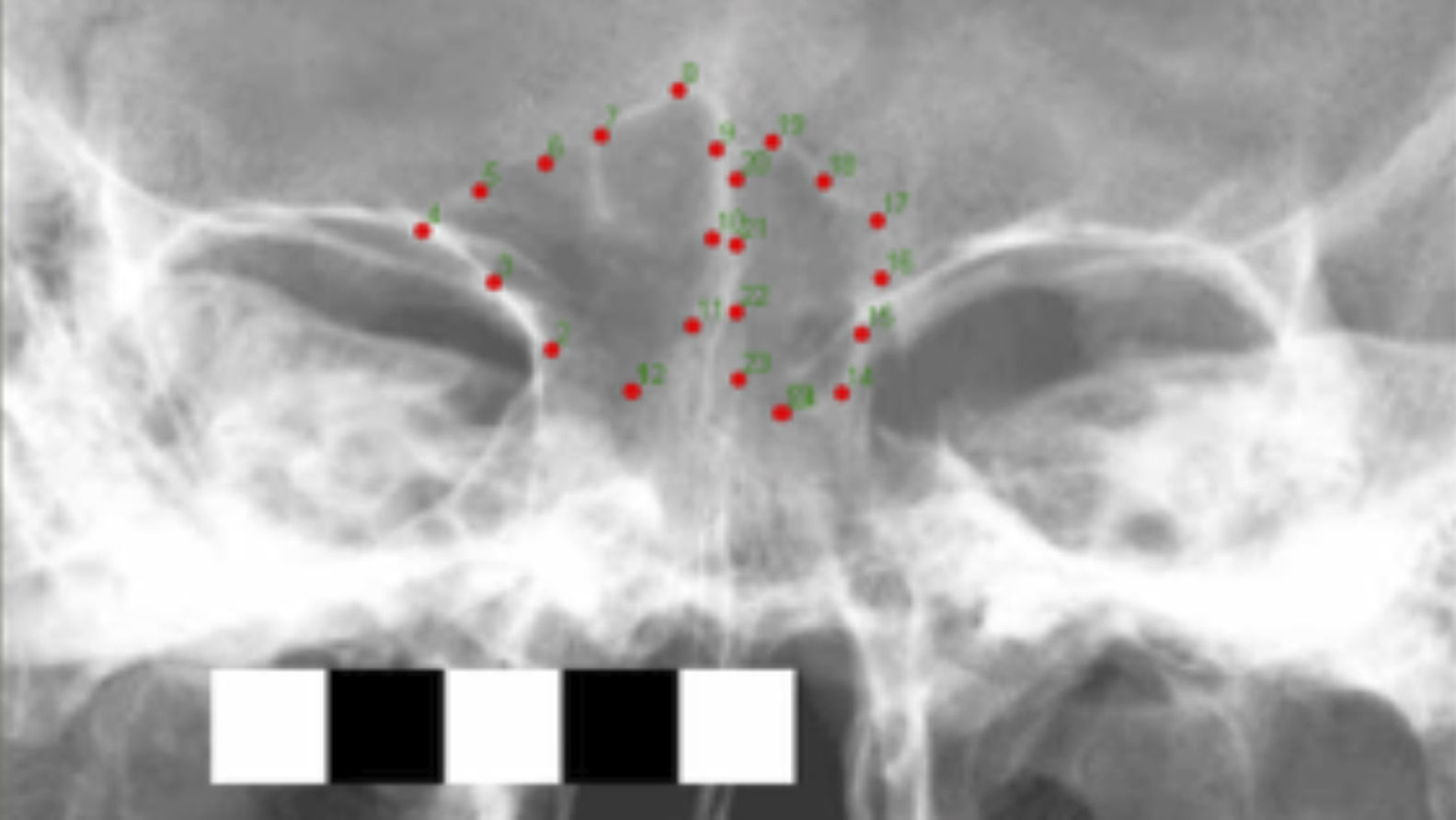
Análisis morfométricosLos análisis morfométricos fueron efectuados sobre imágenes radiográficas y tomográficas del esqueleto facial tomadas en norma frontal con los cráneos posicionados en el plano de Frankfurt. Se digitalizó el contorno de los SF tanto de las radiografías como de las tomografías utilizando un patrón de 24 semilandmarks, el cual se muestra en la figura 1. El registro de las coordenadas de landmarks y semilandmarks se realizó con los programas tpsDig2.13 (Rohlf, 2010), CoordGen6f (Sheets, 2000) y SemiLand6 (Sheets, 2003; Sheets, 2006).

Las coordenadas de landmarks y semilandmarks fueron superpuestas utilizando el criterio del análisis Procrustes generalizado (Rohlf y Slice, 1990). A su vez, los semilandmarks fueron deslizados a lo largo del contorno por medio del criterio de energía de deformación (Bookstein, 1997). Las coordenadas superpuestas fueron empleadas como variables de forma en los análisis estadísticos subsiguientes (Mitteroecker y Gunz, 2009). El programa tpsRelw1.46 (Rohlf, 2010) fue usado para superponer las coordenadas de landmarks y semilandmarks así como también para deslizar las coordenadas de semilandmarks a lo largo del contorno, y finalmente para obtener las variables de forma (i.e., las coordenadas superpuestas).
Con el fin de explorar el patrón de variación en la forma de los SF dentro y entre las muestras se efectuó un análisis de componentes principales (relative warps, RW) sobre las coordenadas superpuestas de los individuos. Los RW describen los ejes de mayor variación en forma en la muestra estudiada y fueron calculados empleando el programa tpsRelw1.46 (Rohlf, 2010).
En el grupo de la Ciudad de México con edad y sexo conocido primeramente se observó el patrón de variación de los SF conforme al incremento de tamaño de los individuos y de la edad cronológica de estos. Seguido a esto, se comparó la forma y el tamaño de los SF masculinos y femeninos en individuos de más de 16años para observar el patrón que existe entre el incremento de edad con el aumento del tamaño y los cambios morfológicos de los SF. Se realizó una regresión lineal entre la edad de los individuos y el tamaño del centroide. También se hizo una regresión lineal entre el logaritmo de la edad de los individuos y el tamaño del centroide. Estos análisis se realizaron con el software Past3.0 (Hammer, Harper y Ryan, 2009). Finalmente, usando el programa Regress6K (Sheets, 2003; Sheets, 2006), se hizo un análisis para establecer la correlación entre las distancias Procrustes de las formas de los SF y el incremento del tamaño con relación a la edad de los individuos.
Para establecer si existen diferencias en el tamaño y la forma de los SF de hombres y mujeres de la Ciudad de México se hicieron pruebas estadísticas con las funciones F de Fisher y t de Student al tamaño del centroide de los SF con el software Past3.0 (Hammer et al., 2009). Después se efectuaron pruebas con la función F en la distancia Procrustes (Zelditch, Swiderski y Sheets, 2012) con 4900 remuestreos para comparar la morfología entre ambos sexos; lo anterior se efectuó con el software TwoGroup6f.
Dado que el objetivo de la investigación es mostrar que el estudio de los SF es fiable para comparar poblaciones, el criterio de inclusión de las muestras únicamente debe cumplir con los requisitos de que los individuos sean mayores de 16años (a esta edad se estima que ya finalizó el crecimiento de los SF) y que no presenten enfermedades que puedan afectar al tamaño y la forma de estas cavidades (Costa, 1978; Rice y Glukman, 1994; Quatrehomme et al., 1996; Kirk et al., 2002; Cameriere et al., 2008).
Finalmente, para comparar los patrones de variación que se presentan entre las poblaciones antiguas de Mesoamérica y la población actual de la Ciudad de México se hizo un análisis de variables canónicas con el programa CVAGen6o (Sheets, 2006). Para comparar la disparidad que presentan las distintas poblaciones analizadas, para este estudio se utilizó el programa DisparityBox6h (Sheets, 2003).
ResultadosEn la muestra de la Ciudad de México no se encontró correlación alguna entre la edad de los individuos y el tamaño de los SF. Al comparar la edad de edad de los individuos con el tamaño del centroide se encontró un coeficiente de correlación no significativo (r=0.43814). Al efectuar el mismo análisis utilizando una transformación logarítmica para la edad de los individuos, nuevamente se obtuvo un coeficiente de correlación no significativo (r=0.44018). Resultados semejantes han sido previamente reportados (Goyal, Acharya, Sattur y Naikmasur, 2013).
En los análisis de regresión entre las distancias Procrustes y la edad de los individuos de la Ciudad de México tampoco se observó correlación alguna. En este caso se obtuvo un coeficiente de correlación entre las variables morfológicas y la edad de r=−0.1303. Se hizo un segundo análisis de regresión, pero esta vez transformando logarítmicamente la edad de los individuos, y el coeficiente de correlación obtenido fue de r=−0.0126. Al comparar el tamaño de los SF entre hombres y mujeres de la Ciudad de México tampoco se encontró una diferencia significativa. Con F=3.0486 (p=0.23798) se comprobó que tanto hombres como mujeres presentan varianzas similares. La prueba de la t de Student t=−1.7814 (p=0.10016) muestra que ambos grupos son estadísticamente similares.
Tampoco se observó una diferencia estadísticamente significativa entre la forma de los SF. El estadístico F Procrustes=2.42 (p=0.05) mostró que no se pueden diferenciar hombres y mujeres de la Ciudad de México basándose en el contorno de los SF.
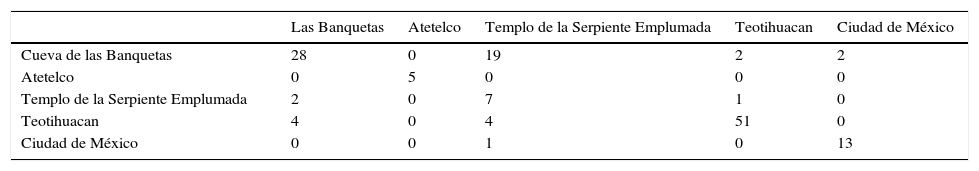
Con esta información se procedió al análisis de las variables canónicas (CVA) para mostrar la variación que presentan las poblaciones mesoamericanas junto con la población de la Ciudad de México. En los cuadros 2 y 3 se presentan los resultados y en la figura 2 los grupos obtenidos a partir de las distancias de Mahalanobis.
Grupos obtenidos a partir del CVA. En las columnas se encuentran los grupos originales y a lo largo de cada una de las filas encontramos los grupos obtenidos tras el análisis de los senos frontales
| Las Banquetas | Atetelco | Templo de la Serpiente Emplumada | Teotihuacan | Ciudad de México | |
|---|---|---|---|---|---|
| Cueva de las Banquetas | 28 | 0 | 19 | 2 | 2 |
| Atetelco | 0 | 5 | 0 | 0 | 0 |
| Templo de la Serpiente Emplumada | 2 | 0 | 7 | 1 | 0 |
| Teotihuacan | 4 | 0 | 4 | 51 | 0 |
| Ciudad de México | 0 | 0 | 1 | 0 | 13 |
Valores de disparidad y error estándar asociados a cada uno de estos valores
| Grupo | Disparidad (diversidad morfológica) | Error estándar | Porcentaje de error |
|---|---|---|---|
| Cueva de las Banquetas | 0.036667 | 0.002345 | 6.4% |
| Atetelco | 0.025018 | 0.004902 | 1.9% |
| Templo de la Serpiente Emplumada | 0.032001 | 0.004500 | 1.4% |
| Teotihuacan | 0.038723 | 0.003084 | 8.0% |
| Ciudad de México | 0.043168 | 0.008158 | 1.8% |

Las poblaciones de Atetelco y la Ciudad de México son las que permanecen mejor agrupadas de acuerdo con las distancias obtenidas con el CVA, ya que un solo individuo de la Ciudad de México se clasifica como perteneciente a la muestra del Templo de la Serpiente Emplumada. En los individuos de Las Banquetas y el Templo de la Serpiente Emplumada observamos que después del análisis de CVA ambos grupos intercambian una gran cantidad de individuos. Algunos individuos de Las Banquetas son clasificados como teotihuacanos o habitantes de la Ciudad de México. Por último, observamos que gran parte de los individuos de Teotihuacan siguen agrupados después del análisis. Sin embargo, de los 59 individuos originales, 4 son clasificados como integrantes del grupo de los sacrificados del Templo de la Serpiente Emplumada y otros cuatro se alinean como parte del grupo de la Cueva de las Banquetas.
En lo tocante a la disparidad, se obtuvo que la muestra con el valor más grande es la de la Ciudad de México, seguida por la de Teotihuacán. La disparidad de cada grupo se presenta en el cuadro 3.
DiscusiónNo se encontró relación entre la edad tamaño y forma de los SF en la población de la Ciudad de México. Asimismo, los resultados muestran que no es posible relacionar el tamaño o la forma de los SF con la edad de los individuos. Tampoco se encontró algún patrón que relacione el tamaño de estos con su forma. Nuestros resultados están de acuerdo con otros estudios que describen los cambios ontogenéticos de los SF como extremadamente variables (Prossinger y Bookstein, 2003).
Tampoco se hallaron diferencias significativas entre la forma o el tamaño de los SF entre hombres y mujeres de la Ciudad de México. En otras poblaciones se ha observado que los hombres poseen SF más grandes (Prossinger, 2001). Una excepción importante se halla en la población esquimal canadiense (Yoshino et al., 1987). También se ha reportado en una muestra moderna de la India que no es factible establecer diferencias en la forma y tamaño de los SF entre hombres y mujeres (Goyal et al., 2013), resultado que se asemeja a los aquí obtenidos con la población mexicana moderna.
Usando el análisis de variables canónicas se observó que la población Posclásica de Atetelco se separa completamente de las poblaciones del periodo Clásico mesoamericano, tanto de Teotihuacan como de Chiapas, y de la población de la Ciudad de México. Las diferencias halladas en la forma y tamaño de los SF con respecto a Atetelco Posclásico, Teotihuacan y el grupo maya se refuerzan con la información arqueológica, ya que Kanjou y Andrade (2003) reportan que los elementos culturales asociados a los entierros son completamente diferentes a los hallados en los contextos del Clásico teotihuacano. Esto está en concordancia con nuestro análisis, llevándonos a inferir que los habitantes del periodo Mazapa no compartían herencia cultural ni biológica con los anteriores pobladores de Teotihuacan.
La población teotihuacana y la población maya de Las Banquetas forman un continuo en la primera variable canónica, encontrándose que la parte negativa es dominada por la población de Teotihuacan y la parte positiva en la población maya. Los individuos encontrados en el Templo de la Serpiente Emplumada presentan una mayor afinidad con las poblaciones mayas que con la población teotihuacana.
En lo referente a la variación que presenta cada población, se observó que la de Ciudad de México es la que presenta una mayor disparidad en la morfología de los SF. Este dato no es sorprendente, ya que la población de Ciudad de México presenta una fuerte inmigración de poblaciones mestizas de todo el país y de poblaciones extranjeras. La segunda población con más variación es la teotihuacana; este dato es concordante con el papel que desempeñó esa urbe como centro económico, teológico y cultural.
De varios años a la fecha, los restos del Templo de la Serpiente Emplumada han generado una serie de interpretaciones sobre su origen, oficio y estatus social. Algunos autores consideran que estos individuos formaban parte del ejército teotihuacano (Cowgill, 1997; White, Spence, Longstaffe, Stuart-Williams y Law, 2002; Spence, White, Longstaffe y Law, 2004), y otros apoyan la tesis de que los sacrificados pudieron ser civiles ataviados con símbolos bélicos con la finalidad de representar el ritual dedicado a la edificación del templo (González Sobrino, 2002; Sugiyama, 2005). También, mediante el uso de técnicas isotópicas, se ha discutido que este grupo de sacrificados son una mezcla de teotihuacanos que habían pasado parte de su vida en diferentes regiones, incluyendo zonas de Guatemala, el Valle de Oaxaca y Michoacán. Se ha reportado que la mayoría de los varones colocados en los entierros periféricos del Templo de la Serpiente Emplumada habían vivido en Teotihuacan antes de morir, mientras que los hombres enterrados debajo de la parte central de la pirámide habían pasado los últimos años antes de morir fuera de Teotihuacan. Las mujeres enterradas en tumbas periféricas parecen haber vivido la mayor parte de sus vidas dentro de Teotihuacán (White et al., 2002; Spence et al., 2004; Nado, Zolotova y Knudson, 2016). Nado et al. (2016) hallaron diferencias en la dieta que sugieren que los seleccionados para el sacrificio de la Pirámide de la Serpiente Emplumada mantuvieron una identidad social o económica distintiva en comparación con el resto de la población teotihuacana, sin especificar alguna identidad étnica ni filiación biológica.
La continuidad que presentan las poblaciones teotihuacanas y mayas, así como su gran disparidad, son indicativos de que debió existir un intercambio genético entre estas poblaciones. Más allá del tema del intercambio comercial y político aún es necesario investigar el tema de la presencia maya en Teotihuacan. Nichols (2016) elabora una interesante discusión al respecto. Cowgill (1997) y Taube (2004) destacan que las relaciones de igualdad o equidad se hicieron más desiguales a medida que Teotihuacan ampliaba sus círculos comerciales. Cowgill (1997) describe un cambio en las interacciones entre los mayas y los teotihuacanos cerca del 350d.C. que involucró intervenciones directas de personas relacionadas con Teotihuacan respaldadas por fuerzas armadas en: Matacapán en la Costa del Golfo, el distrito Cerro Bernal y Mirador en Chiapas, en la costa de Guatemala Tikal, Copan, Kaminaljuyu y tal vez en otros lugares. La Pirámide de la Serpiente Emplumada fue construida durante las fases Miccaotli y Tlamimilolpa temprana (150-250d.C.), período durante el cual Teotihuacan estaba consolidando su influencia en Mesoamérica, y es probable que la selección de las víctimas esté reflejando, además de la parafernalia militar, los cambios en las relaciones con las ciudades mayas y zapotecas. No está de más mencionar que la Cueva de las Banquetas se localiza entre el Altiplano Central y la Depresión Central del actual estado de Chiapas, región intermedia a los emplazamientos identificados con presencia o influencia teotihuacana en los actuales estados de Veracruz, Oaxaca, Tabasco y Centroamérica.
Con nuestros resultados no queremos inferir que los sacrificados de la Pirámide de la Serpiente Emplumada procedan de zona maya. Sin embargo, este es el primer trabajo que discute información morfológica para comparar a los integrantes de este contexto con restos de otros hallazgos teotihuacanos y mesoamericanos. Para tener información más conclusiva, en caso de que se pudiese, sería necesario reintegrar los estudios morfológicos a la discusión de las relaciones interregionales de Teotihuacan. A pesar de que en la actualidad se cuenta con estudios moleculares (Aguirre Samudio et al., 2016; Álvarez-Sandoval et al., 2015; Nado et al., 2016), aún hace falta información concerniente a migraciones y aspectos demográficos entre Teotihuacan y las poblaciones con las que mantenía intercambio comercial y político.
FinanciamientoEste trabajo contó con financiamientos del CONACyT, proyecto “Del símbolo a la herramienta: la industria ósea humana en Teotihuacan”, Apoyo CONACyT, Ciencia Aplicada 100936, y de la DGAPA, UNAM, proyecto PAPIIT IN103815, “Autómatas celulares y selección sexual en primates”.
Se agradece al Mtro. Rubén Cabrera y a los investigadores de la zona arqueológica de Teotihuacan por permitir el estudio de las colecciones óseas.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.