






Durante un período de 4 meses, y mientras se llevaba a cabo un muestreo ambiental de aire, se diagnosticaron 2 casos de aspergilosis por Aspergillus flavus en un centro oncohematológico de Buenos Aires, Argentina.
ObjetivosConocer la variabilidad y la relación genética entre los aislamientos clínicos y los ambientales obtenidos en el centro oncohematológico.
MétodosSe utilizaron 2 técnicas de genotipificación con diferente poder discriminatorio (RAPD y AFLP). Una matriz de similitud genética fue calculada usando el método de Jaccard y fue la base para la construcción de un dendrograma por el método de UPGMA. Se estimó el nivel de variabilidad genética por medio del porcentaje de loci polimórficos, número de alelos efectivos y heterocigosidad esperada, y el índice de asociación (IA).
ResultadosEl dendrograma mostró que los aislamientos de A. flavus recuperados de los pacientes no se relacionaron genéticamente con los del ambiente nosocomial. Los valores más altos de diversidad genética correspondieron a los aislamientos ambientales. El IA estimado para todos los aislamientos sugiere eventos de recombinación.
ConclusionesLos pacientes 1 y 2 no fueron infectados con los aislamientos obtenidos del ambiente hospitalario. Los aislamientos clínicos y ambientales de A. flavus mostraron alta variabilidad genética entre ellos.
During 4 months, and while conducting an environmental sampling of air, 2 cases of aspergillosis by Aspergillus flavus (A. flavus) were diagnosed at an oncohematological center in Buenos Aires, Argentina.
AimsThe aim of this study was to know the variability and the genetic relationship between the clinical and environmental isolates, obtained in the oncohematological center.
MethodsTwo genotyping techniques of different discriminatory power (RAPD and AFLP) were used. A genetic similarity matrix was calculated using Jaccard method and was the basis for the construction of a dendrogram by UPGMA. The level of genetic variability was assessed by measuring the percentage of polymorphic loci, number of effective allele, expected heterocygozity and association index test (IA).
ResultsThe dendrogram reveals that the A. flavus isolates recovered from the patients were not genetically related to those gotten from the rooms occupied by the patients. The environmental isolates had higher values of genetic diversity than the clinical isolates. The IA estimated for all the isolates suggest that recombination events occurred.
ConclusionsPatients 1 and 2 were not infected with isolates from the nosocomial environment. Clinical and environmental isolates of A. flavus showed high genetic variability among them.
Las especies del género Aspergillus están distribuidas ampliamente en el medio ambiente, y cerca de 250 especies se han identificado, de las que más de 40 están documentadas como causantes de enfermedad en el ser humano3. La inhalación de conidios de Aspergillus por el huésped puede inducir manifestaciones alérgicas, colonización, o infecciones invasivas en función de su inmunidad24. Las infecciones por Aspergillus se registran con mayor frecuencia en pacientes inmunocomprometidos y estas son causadas por Aspergillus fumigatus (80%), seguido por Aspergillus flavus (15-20%) y en menor medida por Aspergillus niger y Aspergillus terreus24. En Estados Unidos, A. fumigatus es el agente etiológico más frecuente de la aspergilosis, mientras que A. flavus es el patógeno predominante en países del Medio Oriente, África y el sudeste de Asia, por su habilidad para sobrevivir en condiciones climáticas áridas y secas21,22. A. flavus es una de las especies que tiene afinidad por un amplio rango de huéspedes incluyendo el ser humano, los animales y las plantas, además de ser el patógeno que está involucrado con mayor frecuencia en la producción de aflatoxinas y es capaz de causar otras enfermedades en personas inmunocompetentes tales como sinusitis fúngica indolente, queratitis, otitis y onicomicosis1,7,15.
La tipificación molecular de los aislamientos clínicos y ambientales de A. flavus y A. fumigatus ha proporcionado información importante sobre aspectos epidemiológicos y de salud pública, como son las fuentes de infección, las vías de transmisión, la identificación del patógeno o su resistencia a antifúngicos, lo que sin duda ha coadyuvado a implementar medidas de prevención de la infección en personas susceptibles. Entre las técnicas moleculares que han sido aplicadas en la genotipificación de A. flavus, destaca el empleo de la técnica RFLP (Restriction Fragment Length Polymorphism) para distinguir A. flavus de otras especies relacionadas, así como inferir sus relaciones filogenéticas y discriminar entre aislamientos de la misma o diferentes especies4,6,10,28,33. Otra técnica utilizada ha sido el RAPD (Random-Amplified Polymorphic DNA), que a pesar de presentar problemas en su reproducibilidad, ha permitido la tipificación exitosa de aislamientos clínicos y ambientales de A. flavus, e incluso se ha utilizado para corregir identificaciones equivocadas de aislamientos de otras especies registradas como A. flavus2,5,11,19,34,48. El AFLP (Amplified Fragment Length Polymorphism) también ha sido aplicado en estudios de diversidad y estructura genética de poblaciones de Aspergillus12,32. Otra de las técnicas empleadas es el análisis de microsatélites y la secuenciación de regiones ITS (Internal Transcribed Spacer) o de genes específicos, que son técnicas con mayor poder de discriminación en la tipificación de A. flavus y en la identificación de especies estrechamente relacionadas con este hongo4,5,13,16,25,36,39,40,43,45. Aunque cada una de estas técnicas posee diferentes niveles para la tipificación de microorganismos, actualmente se utilizan 2 o más técnicas moleculares como estrategia para lograr una tipificación más eficiente de los aislamientos de A. flavus y otras especies, como es el caso de la caracterización por multilocus enzyme electrophoresis, RAPD y sequence-specific DNA primers, o RAPD y short sequence repeat16,41. Considerando estos antecedentes, el objetivo del presente trabajo fue genotipificar aislamientos clínicos y ambientales de A. flavus obtenidos en la Institución de diagnóstico y tratamiento de pacientes oncohematológicos de Buenos Aires, Argentina, utilizando el AFLP y RAPD. Además, fue de nuestro interés estimar la diversidad y relación genética entre los aislamientos.
Materiales y métodosAislamientos fúngicosSe utilizaron 6 aislamientos clínicos obtenidos de 2 pacientes oncohematológicos (1 y 2) internados en la Institución de diagnóstico y tratamiento de pacientes oncohematológicos en el período de febrero a mayo de 2005, ambos con diagnóstico de aspergilosis probable según los criterios de consenso de EORTC/MSG8. El paciente 1 estuvo internado en la habitación 23 y el paciente 2 en las habitaciones 10 y 22. Las habitaciones 22 y 23 son contiguas, y ninguna de ellas posee equipamiento de filtración de aire ni presión positiva. Ambas habitaciones comparten un baño externo con ducha, contiguo a la habitación 23, al que acceden los pacientes por el mismo pasillo. El personal de limpieza reporta que toda superficie mojada o húmeda es secada luego de haberse utilizado la ducha.
Los pacientes presentaron, durante su estancia, sinusitis y cuadro neumónico. Cinco aislamientos de A. flavus fueron obtenidos a partir de diferentes muestras clínicas del paciente 1, y un aislamiento a partir de lavado broncoalveolar del paciente 2. Seis aislamientos ambientales de A. flavus fueron recuperados de muestras de aire, tomadas tanto en las habitaciones referidas anteriormente, como en otros sitios dentro y fuera del nombrado nosocomio, entre el 1 de noviembre de 2004 y el 30 de noviembre de 2005. Las muestras de aire se tomaron con el dispositivo portátil SAS Super-90 (Pbi Internacional, Milán, Italia) según las indicaciones del fabricante, a razón de 200l por muestra y descontaminando el equipo con alcohol isopropílico al 70% entre cada toma de muestra. La muestra ambiental del baño compartido por las habitaciones 22 y 23 fue muestreada con idéntica metodología; sin embargo, no se recuperó ningún aislamiento de A. flavus.
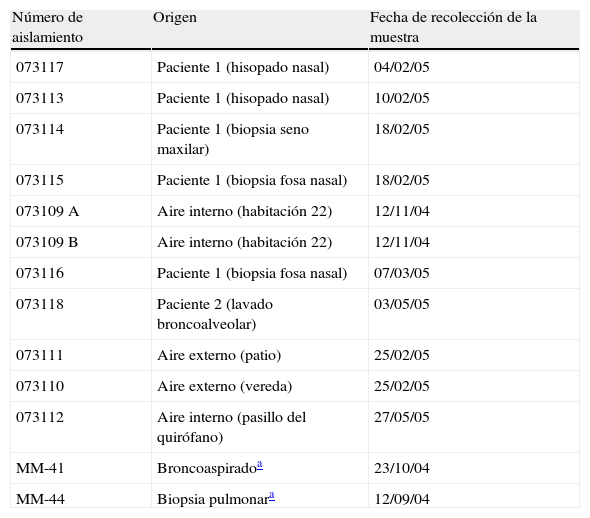
El medio de cultivo utilizado en el muestreo fue agar extracto de malta al 2% con cloranfenicol (250mg/l). Las placas fueron incubadas durante una semana a 28°C con observación diaria. Como testigos externos, se utilizaron 2 aislamientos clínicos de A. flavus provenientes de México. Las características y los datos de recogida de las diferentes muestras y el aislamiento de los hongos se indican en la tabla 1.
Origen y fecha de recolección de las muestras de los aislamientos de A. flavus
| Número de aislamiento | Origen | Fecha de recolección de la muestra |
| 073117 | Paciente 1 (hisopado nasal) | 04/02/05 |
| 073113 | Paciente 1 (hisopado nasal) | 10/02/05 |
| 073114 | Paciente 1 (biopsia seno maxilar) | 18/02/05 |
| 073115 | Paciente 1 (biopsia fosa nasal) | 18/02/05 |
| 073109 A | Aire interno (habitación 22) | 12/11/04 |
| 073109 B | Aire interno (habitación 22) | 12/11/04 |
| 073116 | Paciente 1 (biopsia fosa nasal) | 07/03/05 |
| 073118 | Paciente 2 (lavado broncoalveolar) | 03/05/05 |
| 073111 | Aire externo (patio) | 25/02/05 |
| 073110 | Aire externo (vereda) | 25/02/05 |
| 073112 | Aire interno (pasillo del quirófano) | 27/05/05 |
| MM-41 | Broncoaspiradoa | 23/10/04 |
| MM-44 | Biopsia pulmonara | 12/09/04 |
Paciente 1: internado en la habitación 23 (contigua a la 22). Paciente 2: internado en habitaciones 10 y 22.
Todos los aislamientos fueron identificados por sus características macro y micromorfológicas utilizando las claves de Raper y Fennell37 y Klich y Pitt23. Los aislamientos fueron conservados en agua y medio de cultivo con aceite mineral, y depositados en las colecciones de cultivos fúngicos del Departamento de Micología del INEI-ANLIS Dr. Carlos G. Malbrán y del Laboratorio de Micología Molecular del Departamento de Microbiología y Parasitología de la Facultad de Medicina, UNAM.
Obtención de ADNLos aislamientos fueron cultivados en agar papa dextrosa (Bioxón, México D.F., México) durante 3 días a 28°C. A partir de estos cultivos se obtuvo una suspensión de conidios, agregando a cada tubo 1ml de tampón PBS adicionado con 0,5% de Tween® 20 y agitando en vórtex (Daigger Vortex Genie 2®, Scientific Industries Inc., NY, Estados Unidos). Posteriormente, se sembró en medio líquido YEPD (extracto de levadura 1%, peptona 2%, dextrosa 2%), y se incubó a 37°C con agitación durante 2 días o hasta observar crecimiento micelial. La biomasa micelial de cada aislamiento se filtró al vacío utilizando papel filtro estéril en un embudo Büchner, realizando 2 lavados con agua milliQ® estéril hasta eliminar la humedad.
Posteriormente, la extracción del ADN se llevó a cabo con el Kit DNeasy® Plant Mini (Qiagen, TX, Estados Unidos) de la siguiente manera: 100mg de masa micelial fueron depositados en tubos de 1,5ml conteniendo 0,2g de perlas de vidrio estériles de 400 a 455μm de diámetro (Sigma, MO, Estados Unidos) y se agregaron 400μl del tampón de extracción AP1 (Kit DNeasy® Plant Mini [Qiagen, TX, Estados Unidos]). Los tubos con el micelio se colocaron en un FastPrep®24 (MP Biomedicals, CA, Estados Unidos) y se sometieron a 4 períodos de disrupción celular. Los períodos fueron de 40s a 6m/s, con 5min de enfriamiento en hielo entre cada período. Después de romper el micelio, se continuó con las indicaciones descritas por el fabricante del kit. La concentración de cada muestra de ADN fue cuantificada por fluorometría y por electroforesis en gel de agarosa al 0,8% teñido con bromuro de etidio (10μg/ml), en amortiguador TBE 0.5X (Tris-Base 45mM, ácido bórico 45mM, EDTA 1mM), pH 8,0. El ADN se cuantificó por comparación con diferentes concentraciones del fago λ (10, 20 y 50 ng/μl) (Invitrogen, CA, Estados Unidos). Las imágenes de los geles fueron fotodocumentadas (GeneCam; Syngene, MA, Estados Unidos). Finalmente, el ADN se conservó a 4°C.
Identificación molecular de los aislamientos fenotípicamente identificados como A. flavusLa identidad de los aislamientos de A. flavus incluidos en el estudio fue confirmada por la técnica PCR descrita por González-Salgado et al.14. La PCR se llevó a cabo con pequeñas modificaciones: la mezcla de reacción (25μl) consistió en 5 ng/μl de ADN, dNTPs 200μM, MgCl2 2mM, 100pmol de cada oligonucleótido FLA1 (5’-GTAGGGTTCCTAGCGAGCC-3’) y FLA2 (5’-GGAAAAAGATTGATTTGCGTTC-3’) (Sigma-Aldrich, MO, Estados Unidos) y Taq polimerasa 1U (Invitrogen, CA, Estados Unidos), en amortiguador 1X. Se utilizó como testigo ADN de otros hongos relacionados (A. fumigatus y A. niger) procedentes del cepario del Laboratorio de Micología Molecular del Departamento de Microbiología y Parasitología de la Facultad de Medicina, UNAM. El programa de amplificación fue: un ciclo a 95°C durante 5min; 26 ciclos de 95°C durante 30s, 58°C durante 30s y 72°C durante 45s, y un ciclo de extensión a 72°C durante 5min. Los productos de amplificación se analizaron por electroforesis en gel de agarosa al 1,5%, teñido con bromuro de etidio.
Random-amplified polymorphic DNASe utilizaron 2 oligonucleótidos, 1253 (5’-GTTTCCGCCC-3’) y 1283 (5’-GCGATCCCCA-3’) (Operon Technologies Inc., CA, Estados Unidos), en reacciones independientes. El programa de amplificación, las condiciones de reacción y la lectura de las bandas se realizaron de acuerdo con Reyes-Montes et al.38. Para asegurar la reproducibilidad del RAPD se llevaron a cabo ensayos independientes por duplicado.
Amplified fragment length polymorphismLos análisis de AFLP se llevaron a cabo de acuerdo con lo descrito por Vos et al.46, utilizando el Kit AFLP® Analysis System I (Invitrogen, CA, Estados Unidos). Brevemente, el ADN fue cortado con las endonucleasas EcoRI y MseI. Después de la restricción, los adaptadores fueron ligados a los fragmentos obtenidos. Estos fragmentos fueron preamplificados con los oligonucleótidos E (5’-GACTGCGTACCAATTC-3’) y M (5’-GACGATGAGTCCTGAGTAA-3’), seguido de una segunda PCR selectiva. Los oligonucleótidos selectivos fueron idénticos a los oligonucleótidos E o M, pero con 2 o 3 nucleótidos específicos adicionales en la región terminal 3’. Se utilizaron 6 combinaciones de oligonucleótidos (E+AA/M+CAT; E+AA/M+CAC; E+AC/M+CAT y E+AC/M+CTC; E+AC/M+CAC y E+AA/M+CTC). El oligonucleótido E fue marcado radiactivamente con ATP-P32 y el ADN amplificado fue analizado en geles de poliacrilamida al 5%. La electroforesis se llevó a cabo en una cámara BioRad Laboratories (Sequi-Gen® GT System, CA, Estados Unidos) en un tampón TBE 0.5X, pH 8,0 a voltaje constante (1.600V) y hasta que el colorante xilen-cianol alcanzó el final del gel. Se utilizó el marcador de tamaño molecular DNA Ladder 50bp (Invitrogen, CA, Estados Unidos). El gel se adhirió a papel filtro Whatman no. 1 y se secó al vacío en un sistema HydroTech™ Gel Drying System Model 583 (BioRad, CA, Estados Unidos). Finalmente, se expuso el gel a una placa de radiografía Kodak® X-OMAT AR FILM (35×43cm) a −70°C durante 20h. El análisis de las bandas se realizó en la autorradiografía.
Análisis de datosSe construyó una matriz binaria de datos [presencia (1) y ausencia (0) de bandas] para los 14 aislamientos, a partir de los perfiles obtenidos por AFLP y RAPD. La relación genética entre los aislamientos fue obtenida por medio de un dendrograma construido por el método de UPGMA, a partir de las similitudes genéticas pareadas calculadas con el índice de Jaccard26. La confiabilidad del dendrograma se estableció por medio del coeficiente de correlación cofenética (CCCr), a partir de la prueba aleatorizada no paramétrica de Mantel29. Valores de bootstrap después de 1.000 seudorréplicas fueron obtenidos para conocer la solidez de las agrupaciones en el dendrograma. Estos análisis fueron realizados con los programas NTSYS-PC versión 2.042 y Free Tree35. La diversidad genética de los aislamientos se estimó a partir del índice de Shannon (I), y la heterocigosidad promedio (h), el porcentaje de polimorfismo y el número efectivo de alelos (ne), con el programa POPGENE versión 1.3247. Para distinguir si los aislamientos han experimentado o no eventos de recombinación se utilizó el índice de asociación (IA)30, implementado en el programa LIAN versión 3.518, que mide el grado de asociación no aleatoria entre los alelos de diferentes loci (desequilibrio de ligamiento).
ResultadosTodos los aislamientos de A. flavus incluidos en el estudio presentaron la macro y micromorfología típica descrita para la especie. La identificación se confirmó a través de PCR, donde con los aislamientos de A. flavus se amplificó la banda esperada de 500pb, mientras que los aislamientos de A. fumigatus y A. niger fueron negativos (fig. 1).
: testigo negativo; M: marcador de tamaño molecular DNA Ladder 100bp; MM-41 y MM-44: testigos positivos.")
Identificación molecular de los aislamientos fenotípicamente identificados como A. flavus, con los oligonucleótidos FLA1 y FLA2, específicos para la especie. La electroforesis se llevó a cabo en un gel de agarosa al 1,5% teñido con bromuro de etidio. C(−): testigo negativo; M: marcador de tamaño molecular DNA Ladder 100bp; MM-41 y MM-44: testigos positivos.
El número de bandas obtenidas por RAPD fue de 9 con cada uno de los oligonucleótidos probados y el número de marcadores obtenidos en cada amplificación selectiva del AFLP fue de 69, 60, 100, 97, 35 y 32 para las combinaciones E+AC/M+CAC, E+AA/M+CAC, E+AA/M+CTC, E+AA/M+CAT, E+AC/M+CAT y E+AC/M+CTC, respectivamente. La matriz de datos binarios estuvo integrada por un total de 411 marcadores RAPD y AFLP. La figura 2 muestra los porcentajes de similitud entre los 14 aislamientos estudiados, los cuales se asociaron en 7 grupos. El primero se formó con 4 aislamientos clínicos y 2 ambientales, y este grupo, a su vez, se subdividió en 2 grupos. El subgupo ia incluyó 4 aislamientos, 2 provenientes de diferentes muestras biológicas del paciente 1 (biopsia de seno maxilar e hisopado nasal) y los otros 2 provenientes de muestras ambientales de la habitación 22. El subgrupo ib estaba constituido por 2 aislamientos clínicos del paciente 1 (biopsia de fosa nasal e hisopado nasal). El grupo ii incluyó un aislamiento del ambiente interno de la habitación 10. El grupo iii estaba constituido por un aislamiento del paciente 1 (biopsia de fosa nasal). El grupo iv estaba formado por un aislamiento obtenido del paciente 2 (lavado broncoalveolar). El grupo v estaba formado por 2 aislamientos clínicos de México (broncoaspirado y biopsia pulmonar). El grupo vi, por 2 aislamientos ambientales (vereda y patio del hospital). Finalmente, el grupo vii estaba formado por un aislamiento ambiental (pasillo que conduce al quirófano). Los valores de bootstrap en los diferentes grupos formados presentaron valores superiores al 73% (fig. 2). El valor del CCCr (r=0,95842; p=0,0004) indica que el dendrograma es una buena representación de la relación genética de los aislamientos.
.")
Dendrograma generado por UPGMA, a partir de los perfiles polimórficos obtenidos por RAPD-PCR y AFLP, utilizando el coeficiente de similitud de Jaccard. Los números en los nodos del dendrograma muestran los porcentajes de bootstrap obtenidos después de 1.000 remuestreos (ver apartado de Materiales y métodos).
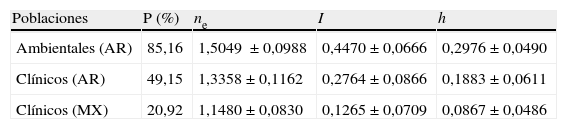
Los diferentes estimadores de variabilidad genética indican que los aislamientos ambientales de A. flavus son más variables que los aislamientos clínicos de Argentina y México. Los estimadores de diversidad genética (P, ne, I, y h) fueron más altos para los aislamientos ambientales que para los clínicos de Argentina y México (tabla 2).
Estimadores de diversidad genética de aislamientos clínicos y ambientales de A. flavus
| Poblaciones | P (%) | ne | I | h |
| Ambientales (AR) | 85,16 | 1,5049 ±0,0988 | 0,4470±0,0666 | 0,2976±0,0490 |
| Clínicos (AR) | 49,15 | 1,3358±0,1162 | 0,2764±0,0866 | 0,1883±0,0611 |
| Clínicos (MX) | 20,92 | 1,1480±0,0830 | 0,1265±0,0709 | 0,0867±0,0486 |
AR: Argentina; h: heterocigosidad; I: índice de diversidad genética de Shannon; MX: México; ne: número efectivo de alelos; P: polimorfismo.
Por último, el IA calculado con todos los aislamientos de A. flavus incluidos en el presente trabajo sugiere que estos han experimentado eventos de recombinación (IA=0,033; p<0,05).
DiscusiónLa aspergilosis nosocomial se asocia con una morbilidad significativa entre los pacientes inmunocomprometidos, lo que tiene implicaciones importantes en el manejo adecuado de los enfermos. Actualmente, A. flavus es la segunda especie más importante del género Aspergillus, causante de infección invasiva, sinusitis y endoftalmitis19,34,44, por lo que existe una necesidad trascendental de llevar a cabo estudios de tipificación molecular para conocer la prevalencia del hongo en ambientes nosocomiales dada su ubicuidad17. Todos los aislamientos estudiados fueron identificados como A. flavus con base en sus características fenotípicas y genotípicas.
Los resultados de la tipificación molecular mostraron que 4 de los 5 aislamientos del paciente 1 están relacionados con los aislamientos de la habitación 22 (grupo i) en un 73% de similitud, mientras que el aislamiento 073116, del mismo paciente, fue incluido en el grupo iii. Este resultado muestra que no se trató de una infección nosocomial, debido a la baja similitud entre estos aislamientos, lo que permite inferir que el paciente 1 tuvo una infección múltiple, probablemente con 5 genotipos diferentes. Asimismo, la ubicación del aislamiento 073118 (paciente 2) en el grupo iv del dendrograma sugiere que el paciente no fue infectado durante su hospitalización, porque su genotipo fue diferente al de los aislamientos ambientales, o bien, que no se aisló el genotipo causante de la infección. Los resultados obtenidos con los aislamientos del paciente 1 apuntan a que un mismo paciente puede albergar diferentes genotipos de A. flavus debido a que el hongo puede sufrir cambios durante su adaptación a los diferentes sitios de colonización43. Al respecto, Leslie et al.27 han propuesto que la duración de la colonización de A. fumigatus en el huésped puede ser un factor importante en la producción y/o selección de cepas, ya que las condiciones microambientales de diferentes sitios anatómicos favorecen o limitan cambios feno y genotípicos en el hongo. También, este tipo de infección por Aspergillus está documentada en enfermos que presentan marcadas alteraciones de las vías respiratorias (fibrosis quística, cavidades pulmonares, bronquiectasias, etc.), por lo que de Valk et al.9 proponen que el tracto respiratorio puede ser colonizado por genotipos diferentes de A. fumigatus, pero que solo uno de ellos puede invadir tejidos adyacentes y subsecuentemente diseminarse a pulmón y otros órganos31.
Por otro lado, los aislamientos obtenidos de las áreas externas del nosocomio y del pasillo del quirófano presentaron baja similitud con los aislamientos de las habitaciones 10 y 22, lo que sugiere que se trata de genotipos diferentes. Estos resultados son apoyados por los estimadores de diversidad genética (tabla 2), el IA y por la presencia de la fase sexual de este hongo20, así como por la fuerte presión selectiva a la que están sometidos los hongos en el medio ambiente.
ConclusionesLos resultados del presente estudio muestran que los pacientes 1 y 2 no se infectaron con los aislamientos del ambiente hospitalario. Sin embargo, la presencia de A. flavus en el ambiente hospitalario debe ser considerada como un factor de riesgo importante para los pacientes.
Declaración de los autoresLos autores declaran que el contenido de este artículo es original y que no ha sido previamente publicado, y que todos los autores han leído y aprobado este manuscrito.
Conflicto de interesesLos autores declaran no tener ningún conflicto de intereses.
El presente trabajo ha sido financiado por la Dirección General de Asuntos del Personal Académico (DGAPA) DGAPA-UNAM-IN224706-3.

