





Los péptidos bioactivos o péptidos con actividad biológica producidos durante la digestión gastrointestinal o la elaboración de alimentos pueden ejercer un importante papel en la regulación y la modulación metabólica, que sugiere su uso potencial como nutracéuticos e ingredientes de alimentos funcionales para promoción de la salud y la reducción del riesgo de enfermedad. En los últimos años se han destinado muchos esfuerzos al estudio de las diferentes actividades beneficiosas que estos péptidos bioactivos pueden tener sobre el organismo, incluyendo su actividad antihipertensiva, hipocolesterolemiante, antioxidante, antimicrobiana e inmunomoduladora, así como su efecto opiáceo. Así mismo se están destinando esfuerzos en investigación para la detección de fuentes alimentarias de péptidos bioactivos así como al estudio de su biodisponibilidad, de sus propiedades funcionales y de sus mecanismos de acción.
Bioactive peptides, or peptides with biological activity produced during gastrointestinal digestion or food processing, could play an important role in metabolic regulation and modulation, suggesting their potential use as nutraceuticals and ingredients of functional foods to promote health and reduce the risk of disease. In the last few years, efforts have been made to study the various potential beneficial activities of these bioactive peptides in the body, including their antihypertensive, hypocholesterolemic, antioxidant, antimicrobial, immunomodulatory, and opiate-like activities. Likewise, research is currently being carried out to detect food sources of bioactive peptides as well as to study their bioavailability, functional properties and mechanisms of action.
Las proteínas que tomamos a través de la dieta nos aportan los aminoácidos necesarios para el desarrollo y el mantenimiento de las células y los tejidos de nuestro organismo. En la actualidad, las proteínas alimentarias se investigan no sólo desde el punto de vista nutricional o funcional, sino como materia prima para la obtención de péptidos1, ya que toda fuente de proteínas alimentarias es susceptible de aportar péptidos funcionales. Sin embargo, hemos de ser críticos en cuanto a su utilización, pese a que en algunos casos su funcionalidad no es algo novedoso, puesto que la existencia de proteínas en la leche materna que contribuyen a la maduración y la función del sistema inmune en el neonato es un hecho conocido desde hace más de 50 años2–4.
Estos péptidos funcionales o bioactivos se definen como secuencias de aminoácidos inactivos en el interior de la proteína precursora, que ejercen determinadas actividades biológicas tras su liberación mediante hidrólisis química o enzimática5. Generalmente, son péptidos de pequeño tamaño, de 3 a 20 aminoácidos, aunque en algunas ocasiones pueden exceder esta longitud, que son liberados durante el procesado industrial de los alimentos, o bien durante la digestión gastrointestinal6,7. Tras la administración oral, los péptidos bioactivos pueden ejercer su efecto, entre otros, sobre los sistemas cardiovascular, digestivo, inmunológico y nervioso8,9. La literatura científica evidencia que estos péptidos pueden atravesar el epitelio intestinal y llegar a tejidos periféricos vía circulación sistémica, pudiendo ejercer funciones específicas a nivel local, en el tracto gastrointestinal y a nivel sistémico. Dentro de estas actividades, los péptidos bioactivos podrían influir en el metabolismo celular y actuar como vasorreguladores, factores de crecimiento, inductores hormonales y neurotransmisores10.
Péptidos bioactivos en alimentosComo se ha comentado anteriormente, los péptidos bioactivos no son más que una pequeña secuencia de aminoácidos encriptados en proteínas; cabe por tanto esperar, debido a la amplia gama de alimentos proteicos existentes en la naturaleza y sobre todo en el mercado, que su ingesta esté asegurada con una alimentación equilibrada. Sin embargo, su biodisponibilidad no está tan clara, ya que han de ser liberados de las proteínas en las que se encuentran tras sufrir la acción de las proteasas gástricas e intestinales y han de poder atravesar el epitelio intestinal y llegar a los tejidos periféricos a través de la circulación sanguínea para poder ejercer su acción9–13.
En los últimos años, debido a la gran relevancia que estos péptidos han obtenido en el mercado, se han desarrollado técnicas para la obtención de nuevos péptidos bioactivos a partir de proteínas alimentarias mediante digestión enzimática in vitro, empleando enzimas proteolíticas de origen microbiano12. Es más, en estudios recientes se han obtenido péptidos modificados, diseñados a partir de péptidos naturales, con el fin de incrementar la actividad de estos últimos14.
De esta manera, proteínas de distinta procedencia (animal y vegetal) han sido utilizadas para el aislamiento de péptidos a partir de hidrolizados enzimáticos9,15,16, siendo interesante destacar que dependiendo de la fuente proteica, del tipo de enzima usado y de las condiciones de procesado, la actividad biológica y los péptidos obtenidos es distinta17. Entre las proteínas de origen vegetal de las que se han aislado péptidos bioactivos encontramos las proteínas de soja, de trigo, de maíz, de arroz, de cebada y de girasol17. En cuanto a las proteínas de origen animal, la leche y otros productos lácteos son los precursores de los péptidos bioactivos más estudiados, aunque también se han identificado estos péptidos en la ovoalbúmina de huevo9, en la carne9, en el músculo de pescado (sardina, atún, bonito)18 y en la jalea real19.
Propiedades beneficiosas de los péptidos bioactivosPropiedades antimicrobianas e inmunomodulantesLa asociación entre la nutrición y la inmunidad es un hecho reconocido desde hace tiempo. Existen estudios que demuestran que péptidos bioactivos derivados de diferentes fuentes de proteínas ejercen efectos inmunomoduladores in vitro e in vivo. Sin embargo, la mayoría de los estudios se centran en la evaluación del efecto de los péptidos e hidrolizados de proteínas específicas en el sistema inmunológico y sólo un número limitado de investigaciones examinan su impacto en la inmunidad inespecífica (innata). Los péptidos inmunomoduladores son liberados de las proteínas que los contienen de manera natural durante el proceso de la digestión en el tracto gastrointestinal, lo que afecta a la respuesta inmunológica y a la función celular en el mismo.
Los péptidos bioactivos con propiedades antimicrobianas e inmunomoduladoras más estudiados son aquellos que proceden de la leche y los productos lácteos20. Este efecto parece estar relacionado con la carga neta positiva de estos péptidos, que se organizan estructuralmente y provocan la formación de canales iónicos en la membrana de los microorganismos21, alterando su permeabilidad y provocando la muerte celular22.
En la leche y también en el huevo se encuentran la lactotransferrina y la ovotransferrina. Ambas transferrinas poseen actividad antibacteriana frente a gran variedad de microorganismos. Esta actividad puede ser ejercida al menos mediante tres mecanismos distintos23–25:
- •
Secuestrando el hierro e impidiendo su utilización por las bacterias.
- •
Produciendo alteraciones en la pared bacteriana.
- •
Mediante la estimulación de la fagocitosis por macrófagos y monocitos.
Los péptidos bioactivos con actividad antimicrobiana ejercen un efecto inhibidor sobre los microorganismos de destino también mediante la interacción con los componentes intracelulares aniónicos como el ADN y el ARN, lo que inhibe la síntesis de proteínas y la división celular de los microorganismos. Por otra parte, algunos péptidos están involucrados en la activación autolítica en los microorganismos diana26.
Péptidos derivados de las caseínas de la leche muestran también actividad antimicrobiana in vitro frente a una amplio rango de microorganismos, entre los que se incluyen Staphylococcus spp y Streptococcus pyogenes. Estudios in vivo muestran efectos protectores de la israicina (derivado de la caseína) frente a S. aureus y Candida albicans en ratones y frente a mastitis en vacas y ovejas27,28.
También se ha descrito que la lactoferrina es capaz de inhibir la replicación de virus como el de la leucemia de células T del tipo I, el citomegalovirus, el virus de la hepatitis C o el herpes simple tipo I14.
La lisozima, presente también en la leche y el huevo, posee actividad antimicrobiana e inmunorreguladora, pudiendo mejorar la sinusitis crónica y normalizar la respuesta humoral y celular en pacientes con bronquitis crónica cuando se combina con inmunoterapia. Además, mejora la respuesta inmune en pacientes cancerosos inmunodeprimidos. Se ha sugerido que la inmunomodulación producida por la lisozima puede ser resultado de la estimulación de la función fagocítica y de la hidrólisis de productos de peptidoglucano que pueden actuar como adyuvantes o inmunomoduladores25,29–31.
Otros alimentos, aunque no tan estudiados como los lácteos, también contienen péptidos con actividad inmunomodulante, como es el caso de la jalea real de la que se han aislado péptidos que estimulan la proliferación de monocitos humanos32, péptidos derivados de las proteínas del arroz y la soja que estimulan los sistemas de inmunidad no específica33 y péptidos del trigo sarraceno y pimiento34,35.
Actividad opiáceaLos péptidos con actividad opiácea, también llamados exorfinas, se definen como péptidos que presentan afinidad por receptores opiáceos y actúan, mediante la unión a receptores, como moduladores exógenos de la motilidad intestinal, de la permeabilidad epitelial y de la liberación de hormonas. Estos péptidos han sido aislados de cereales, leches y tejidos animales11.
La péptidos opioides más estudiados son los derivados de proteínas lácteas (caseínas)11,14. Una vez absorbidos, estos péptidos pueden viajar al cerebro y otros órganos y ejercer actividades farmacológicas similares a la morfina o el opio. Éste puede ser el motivo por el cual los recién nacidos humanos, en general, se calman y tienen sueño después de beber leche. En contraste con las casomorfinas, algunos péptidos producidos por el desglose de κ-caseína pueden actuar como antagonistas de los opioides, es decir, que pueden inhibir el efecto de sustancias como la morfina36.
En los seres humanos adultos, los efectos de estos péptidos suelen limitarse al tracto gastrointestinal, ya que, o bien no son absorbidos, o escapan a la degradación enzimática en la pared intestinal37. Sin embargo, existen estudios que sugieren que algunos de ellos (β-casomorfinas) pueden ser transportados de la sangre al tronco cerebral38 y al sistema cardiovascular en los lactantes37.
Los péptidos lácteos pueden participar en la regulación del apetito, modificando la actividad endocrina del páncreas para aumentar la producción de insulina39. Estudios en ratas indican que las β-casomorfinas pueden modular la ingesta, estimulando la ingesta de grasa y reduciendo la ingesta de dietas ricas en hidratos de carbono40. En este sentido, también algunos péptidos derivados del gluten de trigo administrados por vía intravenosa y oral en ratas dio lugar a la estimulación postprandial de la liberación de insulina41.
Parte de la función de las casomorfinas podría ser la reducción de la motilidad intestinal, lo que permitiría la máxima producción de otros péptidos bioactivos42. En humanos adultos se han detectado fragmentos opiáceos en el contenido intestinal tras la ingestión de leche de vaca43, no detectándose, sin embargo, en el suero sanguíneo. La confirmación en el hombre de los efectos detectados en los animales de experimentación abriría un nuevo campo de aplicación de estos péptidos opiáceos en la prevención y el tratamiento de trastornos intestinales.
Actividad antioxidanteLos antioxidantes no sólo son importantes en la prevención de la oxidación en los alimentos, sino también a nivel fisiológico. Los radicales libres son unas moléculas que están en continua formación en las células y que son neutralizados de manera natural por las defensas antioxidantes del organismo. Sin embargo, existen factores que producen un desequilibrio en el número de radicales libres, y un exceso de éstos puede dar lugar a un envejecimiento celular o al desarrollo de enfermedades44. Todo esto ha llevado a la búsqueda de compuestos antioxidantes naturales para la prevención de la oxidación por parte de los radicales libres.
El estudio de péptidos antioxidantes presentes en alimentos ha sido un reto en los últimos años, habiéndose identificado numerosos péptidos que presentan actividad antioxidante en diferentes alimentos45,46 y en derivados de ellos, como hidrolizados de gelatina47, hidrolizados de clara de huevo48 y productos lácteos49, entre otros. Estos péptidos actúan impidiendo que otras moléculas se unan al oxígeno, al reaccionar-interactuar más rápido con los radicales libres que con el resto de las moléculas presentes en un determinado microambiente de membrana plasmática, citosol, núcleo o líquido extracelular50.
Actividad antihipercolesterolémicaLa hiperlipidemia, en especial la hipercolesterolemia, es uno de los más importantes factores de riesgo que contribuyen al desarrollo de enfermedades cardiovasculares.
Recientemente se ha demostrado que los péptidos de la soja pueden ser responsables, al menos en parte, de su actividad hipocolesterolémica, ya que se ha observado que hidrolizados de proteína de soja mostraron un mayor descenso del colesterol plasmático que la proteína de soja intacta51.
Este efecto hipocolesterolemiante de los péptidos bioactivos se atribuye a dos acciones de los mismos:
- •
Los péptidos bioactivos inhiben la absorción del colesterol, posiblemente debido a la represión de la solubilidad micelar del colesterol52.
- •
Algunos péptidos pueden regular al alza los receptores LDL, que están crónicamente suprimidos por la hipercolesterolemia o administración de colesterol de la dieta53.
Además de los péptidos derivados de la soja, otros péptidos bioactivos con efecto hipocolesterolémico han sido obtenidos a partir de B-Lactoglobulina, hidrolizado de proteína de carne de cerdo54,55 y proteína vegetal (Brassica carinata)56.
Péptidos con actividad sobre el sistema cardiovascularLos principales efectos descritos de los péptidos bioactivos sobre el sistema cardiovascular son los relativos a su actividad antitrombótica y antihipertensiva.
Actividad antitrombóticaLa agregación plaquetaria es un paso crítico en la formación de trombos y está mediada por el enlace del fibrinógeno y su receptor, la glucoproteína GPIIb/IIIa de la membrana de las plaquetas, la cual involucra el reconocimiento de la secuencia Arg-Gli-Asp (RGD) del fibrinógeno. Es concebible, por lo tanto, que los péptidos que contienen la secuencia RGD puedan ser capaces de antagonizar el acoplamiento del fibrinógeno a la GPIIb/IIIa, resultando en la inhibición de la agregación de plaquetas. En la secuencia RGD, una unidad catiónica, en este caso el grupo guanidino de la cadena lateral de la Arg y la estructura del ácido β-carboxílico de la Asp, es requerida para la actividad inhibitoria. Además, la distancia entre estos grupos funcionales catiónicos y aniónicos es un factor importante en potencia. El fibrinógeno enlaza sitios de la GPIIb/IIIa, la cual tiene un número de residuos de Asp, y a causa de esto el grupo guanidino de la Arg, en el caso de compuesto tipo-RGD, se piensa que puede estar involucrado en la ligadura iónica con el grupo carboxilato del Asp en la GPIIb/IIIa57. Por lo anterior, los péptidos que son antagonistas del fibrinógeno y se fijan a GPIIb/IIIa son útiles en la prevención de la trombosis y en los regímenes de tratamiento post-angioplastia o post-trombolíticos.
Los principales péptidos aislados con actividad antitrombótica se encuentran presentes en la leche. A nivel molecular, la coagulación de la sangre y de la leche muestra gran similitud. En la leche, el mecanismo de coagulación es definido por la interacción de la k-caseína con quimosina, y el proceso de coagulación de la sangre es definido por la interacción del fibrinógeno con trombina.
Determinadas secuencias peptídicas de la κ-CN, denominadas casoplatelinas, son similares a la cadena γ del fibrinógeno58. Se ha comprobado in vitro que los péptidos derivados de la κ-CN bovina son inhibidores de la agregación plaquetaria y de la unión de la cadena γ del fibrinógeno humano al receptor específico de la membrana de las plaquetas59. Estos péptidos son capaces de fijarse sobre receptores específicos situados en la superficie de las plaquetas, impidiendo así la formación del trombo.
Péptidos derivados de la lactoferrina presentan también actividad antitrombótica a través de otro mecanismo distinto al de la caseína, inhibiendo, de manera dosis-dependiente, la agregación plaquetaria inducida por ADP, debido a la homología con el fragmento f(572-575) de la cadena α del fibrinógeno60. Se ha comprobado in vivo que este fragmento inhibe la formación de trombos en la arteria coronaria de perros61 y prolonga el tiempo de sangrado en la arteria mesentérica del hámster62.
En el plasma de recién nacidos alimentados con leche materna o con fórmulas infantiles elaboradas con leche de vaca se han encontrado secuencias peptídicas en concentración suficiente para ejercer el efecto antitrombótico in vivo, lo que demuestra su liberación a partir de las proteínas lácteas durante el proceso de digestión gastrointestinal63. Estudios más recientes han demostrado la presencia de péptidos antitrombóticos en el estómago, duodeno y posteriormente en la sangre de adultos que habían ingerido leche o yogur63.
Actividad antihipertensivaLa hipertensión es la principal causa de enfermedad en los países industrializados. El 35% de la mortalidad en estos países se debe a esta enfermedad o a alguna de sus complicaciones renales, cardiacas o cerebrales. Debido a esto, los alimentos que presentan péptidos con actividad antihipertensiva son de gran interés.
El mecanismo antihipertensivo más estudiado como actividad de los péptidos bioactivos es la inhibición de la actividad de la enzima convertidora de angiotensina (ECA). La ECA es una enzima que cataliza la conversión de la angiotensina I (decapéptido) en angiotensina II (octapéptido).
La angiotensina II es un compuesto de elevada potencia vasoconstrictora. Su acción provoca la contracción rápida de las arteriolas y, por tanto, el incremento de la presión arterial, ya que estimula la secreción de aldosterona por las glándulas suprarrenales, hormona que induce la excreción de potasio y la retención de sodio y agua y provoca el incremento del volumen extracelular, y la neutralización de la producción de renina. La renina libera el decapéptido angiotensina I del sistema renina-angiotensina. Este sistema es, quizás, el más importante de los diferentes mecanismos vasoconstrictores y vasodilatadores implicados en la regulación de la presión sanguínea64.
La ECA actúa simultáneamente en el sistema cinina-calicreína catalizando la degradación de las bradicininas, compuestos de potente acción vasodilatadora, y de esta manera se favorece el incremento de la presión arterial. El incremento de la disponibilidad de bradicininas debido a la inhibición de la ECA puede ser parcialmente responsable del efecto beneficioso de la inhibición de la ECA sobre la presión sanguínea.
Algunos trabajos se han centrado en la producción y el aislamiento de péptidos inhibidores de la ECA, procedentes de proteínas de diferentes alimentos65–67. Algunos autores han encontrado que, en productos fermentados, péptidos con actividad inhibitoria de la ECA podrían, también, presentar actividad antioxidante, sugiriendo la presencia de actividad multifuncional en estos compuestos68,69. Se han buscado inhibidores competitivos exógenos de esta enzima, a partir de distintas fuentes (pescado, maíz, gelatina, veneno de serpiente, vino, etc.). El origen de estas fuentes proteicas engloba tanto al reino vegetal como al reino animal. Los inhibidores competitivos compiten con el sustrato por el centro activo de la enzima. En este caso, el aumento en la concentración del sustrato da lugar a una disminución de la capacidad del inhibidor para competir con éste. Por otra parte, los inhibidores no competitivos se unen a la enzima en una zona diferente al centro activo, deformándolo, de manera que impide el enlace con el sustrato, siendo este tipo de inhibición independiente de la concentración de sustrato. Dependiendo del tipo de inhibidor, se puede producir inhibición competitiva o no competitiva65.
Se han identificado varios péptidos endógenos que actúan como inhibidores y sustratos competitivos de la ECA, como las encefalinas, las bradicininas y la sustancia P. Como primeros inhibidores exógenos de la ECA se estudiaron los extractos del veneno de la serpiente Bothrops jararaca. Actualmente, un derivado sintético, el captopril (D-3-mercapto-2-metilpropanoil-1-prolina; IC50=0,006μM), obtenido tras el estudio del modelo hipotético del sitio activo de la enzima, es el fármaco más empleado en el control de la hipertensión70.
Otra de las enzimas involucradas en la regulación de la presión arterial es la enzima convertidora de endotelina (ECE). Esta enzima cataliza la formación de endotelina-1 a partir de su precursor. La endotelina-1 es un potente vasoconstrictor, lo que favorece el incremento de la presión arterial; por lo tanto, la inhibición de la ECE produce un efecto antihipertensivo. Sólo se han descrito péptidos inhibidores de esta enzima a partir de un hidrolizado digestivo del bonito71, pero el alto contenido y la gran variedad de proteínas presentes en la leche podrían abrir un nuevo campo de estudio encaminado a la identificación de péptidos lácteos con esta actividad.
Durante los últimos 20 años, muchos péptidos inhibidores de la ECA con propiedades antihipertensivas in vivo se han identificado en alimentos.
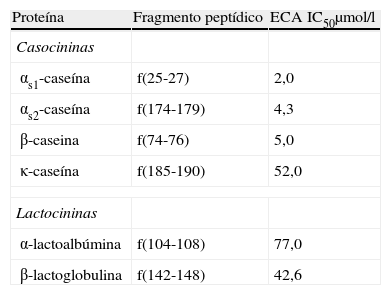
Péptidos inhibidores de ECA en lácteosUn gran número de péptidos inhibidores de la ECA se han aislado de productos lácteos (queso, leche, leche fermentada…). Algunos de ellos han mostrado efectos antihipertensivos en animales de experimentación espontáneamente hipertensos y en pacientes con hipertensión. En la tabla 1 se muestra la actividad antihipertensiva de distintas proteínas lácteas.
Actividad inhibidora de ECA de proteínas lácteas
| Proteína | Fragmento peptídico | ECA IC50μmol/l |
| Casocininas | ||
| αs1-caseína | f(25-27) | 2,0 |
| αs2-caseína | f(174-179) | 4,3 |
| β-caseina | f(74-76) | 5,0 |
| κ-caseína | f(185-190) | 52,0 |
| Lactocininas | ||
| α-lactoalbúmina | f(104-108) | 77,0 |
| β-lactoglobulina | f(142-148) | 42,6 |
Fuente: FitzGerald et al76
ECA IC50: concentración de péptido que inhibe la actividad de la enzima convertidora de angiotensina al 50%.
Maruyama y Suzuki72 fueron los primeros en aislar un péptido inhibidor de la ECA (llamado CEI 12) a partir de un hidrolizado de caseína (dodecapeptido, IC50=77μ M). Posteriormente, otros péptidos inhibidores de la ECA se han aislado de diferentes productos lácteos, sobre todo de productos lácteos fermentados.
Yamamoto et al73 han demostrado que las leches fermentadas con L. helveticus son las que más actividad inhibitoria de la ECA presentan, debido a la alta actividad proteolítica extracelular de esta cepa. Otros péptidos con actividades inhibidoras se han identificado en leches fermentadas por Lactobacillus delbrueckii sp. bulgaricus o Lactobacillus lactis sp. cremoris74 y en el kéfir, una leche fermentada preparada a partir de un caldo de cultivo que incluye bacterias acidolácticas, una cepa de Saccharomyces kefir y algunas especies bacterias acéticas, aunque en éste la actividad inhibidora de la ECA es baja75. Parece que los péptidos que se encuentran en el kéfir no son las principales sustancias involucradas en sus efectos antihipertensivos.
Los estudios llevados a cabo en quesos han demostrado una fuerte actividad inhibitoria de la ECA observada después de un proceso de digestión proteásica76. La proteólisis que tiene lugar durante la maduración de los quesos podría aumentar la tasa de actividad inhibitoria de la ECA a un cierto nivel, más allá del cual esta actividad inhibitoria disminuye77 debido a que los péptidos bioactivos se transforman en moléculas inactivas y aminoácidos78. El momento óptimo de maduración para la producción de péptidos bioactivos es distinto para cada tipo de queso. Como ejemplo, podríamos hablar de 8 meses para un queso tipo manchego79.
En la tabla 280 aparecen reflejados algunos de los productos que existen en el mercado y que ofrecen alegaciones acerca de la reducción de la presión arterial en humanos, las dosis utilizadas y la presión arterial inicial de los sujetos en los estudios.
Estudios clínicos con productos lácteos y péptidos que presentan un efecto antihipertensivo en humanos
| Producto | Componente activo (dosis) | Tensión arterial inicial de los sujetos | Efecto en la presión arterial sistólica |
| Calpis (leche fermentada) | IPP (1,04 mg)VPP (1,42 mg) | Hipertensos | –9,4 mmHg (4 semanas)–14,1 mmHg (8 semanas) |
| Calpis (leche fermentada) | IPP (1,15 mg)VPP (1,98 mg) | Hipertensión media | –4,3 mmHg (2 semanas)–5,2 mmHg (4 semanas) |
| Péptido ameal (hidrolizado de caseínas) | IPP+VPP (1,8 mg)IPP+VPP (2,5 mg)IPP+VPP (3,6 mg) | Presión arterial normal altaHipertensión moderada | –6,3 mmHg (6 semanas)–6,7 mmHg (6 semanas)–10,1 mmHg (6 semanas) |
| Evolus/Kaiku Vitabrand (leche fermentada) | IPP (2,25 mg)VPP (3-3,75 mg) | Hipertensión moderada | –14,9 mmHg (8 semanas) |
| Evolus/Kaiku Vitabrand (leche fermentada | IPP (2,25 mg)VPP (3-3,75 mg) | Hipertensos | –6,7 mmHg (21 semanas) |
| Evolus/Kaiku Vitabrand (leche fermentada) | IPP (2,4-2,7 mg)VPP (2,4-2,7 mg) | Hipertensión moderada | –16 mmHg (8-10 semanas)–11 mmHg (5-7semanas) |
| Biozate 1 (hidrolizado de proteínas de suero) | 20 g de hidrolizado | Hipertensos | –11 mmHg (-7mmHg DBP) comparado con el control. |
Fuente: López Fandiño77.
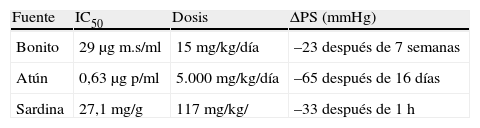
Algunos péptidos inhibidores de la ECA han sido obtenidos por digestión proteica enzimática a partir de proteína muscular de atún81, músculo de sardina82, bonito seco83, algas84 y otros productos marinos.
Algunos hidrolizados de proteínas han mostrado efectos in vitro (mediante la inhibición de la ECA) e in vivo (efectos antihipertensivos) (tabla 3). La administración de un hidrolizado de músculos de bonito a ratas espontáneamente hipertensas ha permitido suprimir la hipertensión después de un periodo de cinco semanas de tratamiento utilizando una dosis de 15 a 60mg/kg60.
Los hidrolizados de algas son también una fuente importante de péptidos biológicamente activos, algunos de los cuales han sido identificados y presentan las actividades inhibitorias de la ECA (tabla 3). Estos resultados pueden explicar en parte los efectos antihipertensivos observados en ratas espontáneamente hipertensas (SHR)83,85. Otros péptidos inhibidores de la ECA han sido identificados en hidrolizados de pescado, pero muy pocas han sido evaluadas in vivo.
Productos vegetalesSe han encontrado péptidos inhibidores de la ECA en muchas fuentes vegetales, como soja9, germen de trigo86 y garbanzo87.
En el gluten de trigo se aisló un hexapéptido que mostró una actividad antagónica con respecto a la angiotensina I in vivo. Recientemente, un hidrolizado de gluten ha conseguido una reducción de la presión arterial en ratas espontáneamente hipertensas 2h después de la administración intravenosa88.
La hidrólisis de las proteínas de colza reveló que la presencia de péptidos inhibidores de la ECA puede mostrar un efecto antihipertensivo in vivo. Este hidrolizado, administrado a ratas SHR, reduce significativamente la presión arterial 2h después de su administración por vía oral (0,15g/kg)89. También se ha demostrado el efecto antihipertensivo de un régimen de soja cuando se administra en ratas SHR90. De la misma manera, la digestión de las principales proteínas de reserva de semillas de garbanzo produce péptidos inhibidores de la ECA91.
Otras fuentesOshima et al91 fueron los primeros en identificar péptidos inhibidores de la ECA a partir de proteínas de la dieta. En los últimos años se han llevado a cabo estudios que describen la existencia de péptidos con actividad antihipertensiva en otras fuentes alimentarias, como músculo de cerdo y pollo, huevos, ajo, gelatina, sake, vino, autolisados bacterianos, hígado, calamar, hongos, jalea real y algunas frutas y tés11,92. Sin embargo, la información sobre péptidos bioactivos generados a partir de las proteínas de estos alimentos es todavía limitada.
En el momento actual se han descrito algunos péptidos con actividad vasodilatadora e inhibidora de la ECA, que provienen de la ovoalbúmina hidrolizada con diferentes enzimas. Por ejemplo, la administración crónica (12 semanas) a ratas SHR de un hidrolizado de yemas de huevo ha eliminado el desarrollo de la hipertensión en dosis de 20, 100 y 500mg/kg administrados diariamente por vía oral. Dosis más altas administradas a animales han disminuido alrededor del 10% las presiones sistólica y diastólica medias, en comparación con las de los animales testigos93.
Biodisponibilidad de los péptidos bioactivosEl potencial efecto hipotensivo de los péptidos inhibidores de la ECA depende de su capacidad para alcanzar intactos los órganos donde van a realizar su función. Como ya se ha comentado, son muchos los estudios realizados en los que se evalúa la actividad IECA de estos péptidos, pero es difícil establecer una relación directa entre la capacidad de inhibir la ECA in vitro y la actividad antihipertensiva in vivo. El estudio de la biodisponibilidad de los péptidos bioactivos es fundamental a la hora de diseñar alimentos funcionales con propiedades fisiológicas94. Se sabe que dipéptidos y tripéptidos pueden transportarse a través de la mucosa intestinal, aunque también hay evidencias de que péptidos de mayor tamaño pueden absorberse en el intestino delgado63. Se ha demostrado, por tanto, que algunos péptidos con actividad IECA y antihipertensiva son capaces de absorberse en el tracto gastrointestinal acumulándose en diversos órganos, por lo que podrían ejercer su mecanismo de acción sistémicamente19. Sin embargo, se postula que determinados péptidos antihipertensivos podrían no absorberse en el tracto intestinal, ejerciendo una función directa en el lumen intestinal, o bien podrían ejercer su función mediante la interacción con receptores que se encuentran en la pared del intestino.
Una vez absorbidos, el elevado número de peptidasas que se encuentran en la sangre hace que el tiempo durante el que ciertos péptidos permanecen estables en el plasma sea muy corto. Sin embargo, existen algunas evidencias de que péptidos con actividad IECA pueden absorberse en el tracto digestivo y ser transportados por el torrente circulatorio sin ser hidrolizados por las proteasas sanguíneas13.