



BASICA
Cómoactúa el alcohol en nuestro cerebro
Mechanismsof ethanol actions on the brain
GUERRI, C.
Instituto de Investigaciones Citológicas
Fundación Valenciana de Investigaciones Biomédicas.Valencia.
Correspondencia:
CONSUELO GUERRI.
Instituto de Investigaciones Citológicas.
Amadeo de Saboya, 4.
46010-Valencia.
e-mail: guerri@ochoa.fib.es
Resumen: Objetivo: se revisan los efectos del etanol en cerebro, losmecanismos que participan en la tolerancia y dependencia asícomo las consecuencias neuropatológicas a que conduce elabuso de alcohol.
Material ymétodos: se describen los datos existentes sobre losefectos del etanol en cerebro, dando un especial énfasis alas nuevas hipótesis basadas en trabajos recientes enbiología celular y molecular.
Resultados: los efectos eufóricos, hipnóticos yde refuerzo del etanol, parecen estar mediados por unaactivación en la neurotransmisión inhibitoria,vía receptor del ácido*-aminobutírico(GABAA), por una inhibición de losreceptores para glutamato, N-metil-D-aspartato (NMDA) y por unaactivación del sistema dopaminérgicomesolímbico. Otros sistemas de neurotransmisión y elacetaldehído también pueden participar en laacción del etanol en cerebro. El consumo crónico dealcohol induce cambios neuroadaptativos (tolerancia), aumentando elnúmero de los receptores para glutamato NMDA,desensibilizando la respuesta y el número de receptoresGABAA. Las manifestaciones del síndrome deabstinencia parecen estar mediadas por un aumento en laneurotransmisión excitatoria con respecto a la inhibitoria,mientras que en el deseo por reiniciar el consumo de alcohol puedeestar involucrada una reducción en la actividaddopaminérgica mesolímbica. La hiperexcitabilidad enel síndrome de abstinencia puede contribuir al dañocerebral inducido por el etanol.
Conclusiones: la multitud de nuevos datos está ayudandoa esclarecer los mecanismos involucrados en la acción deletanol, tanto en la intoxicación aguda como en lacrónica. Los nuevos hallazgos están conduciendo aldesarrollo de nuevos tratamientos para elalcoholismo.
PalabrasClave: Etanol. Tolerancia. Dependencia. Síndrome deabstinencia.
Abstract: Objective: our knowledge on the targets of ethanol actions onthe brain and on the mechanisms of ethanol tolerance and dependenceis reviewed. The connsequences of alcohol abuse will also bediscussed
Materialand methods: we review existing data on the effects of ethanolon the central nervous system, giving special emphasis to newhypotheses based on recent work on cellular and molecularbiology.
Results: the euphoric, hipnotic and reinforcing effects ofethanol appear due to enhanced inhibition mediated by*-aminobutyric acidreceptors (GABAA), to decreased excitation via inhibitionof N-methyl-D-aspartate (NMDA) glutamate receptors, and toactivation of the mesolimbic dopaminergic system. Otherneurotransmitter systems and acetaldehyde might also participate inethanol´s actions. Tolerance to chronic ethanol consumptionappears due to up-regulation of NMDA glutamate receptors anddesensitization and/or down-regulation of GABAAreceptors. The withdrawal manifestation of alcohol dependence mayresult from enhancement and decrease of excitatory and inhibitoryneurotransmission, respectively, whereas the compulsion toreinitiate alcohol drinking may involve reduction in the mesolimbicdopaminergic activity. Hiperexcitability neurotransmissioncontribute to alcohol-induced brain damage.
Conclusions: a pletora of new data is helping to clarify theprecise mechanisms involved in the acute and chronic effects ofalcohol. This knowledge is guiding the development of newtreatments for alcoholism.
Key Words:Ethanol. Tolerance. Dependence. With drawal.
Introducción
El uso y abusode bebidas alcohólicas y sus propiedades eufóricas,desinhibitorias y depresoras del sistema nervioso central se conocedesde hace siglos y en algunos casos, está asociada aciertas tradiciones, leyendas y leyes de diferentes culturasincluyendo la egipcia, griega y romana. Así, los griegos enla Ilíada, ya describen la costumbre de dar vino a lossoldados antes de entrar en combate, posiblemente para inhibir elmiedo a la batalla.
De hecho,actualmente sabemos que el alcohol es una droga psicotropa,depresora del sistema nervioso central (SNC), con acciónreforzante positiva y con capacidad de crear tolerancia ydependencia psicofísica y adicción. La accióndepresora del etanol sobre el SNC y su capacidad para afectar lasfunciones cognitivas, perceptivas y reducir las capacidadesmotoras, hace que su consumo esté relacionado con aumentosen accidentes laborales y de circulación. Además, adiferencia de otras sustancias adictivas, el etanol es untóxico celular, lo que conlleva a que su consumo ocasionegraves alteraciones en un gran número de tejidos y sistemasdel organismo, con el desarrollo de las consiguientes enfermedadescrónicas, incremento en la mortalidad y elevación delos costes sanitarios.
La simplicidadde la molécula del etanol y la gran diversidad de efectosque produce, ha conducido al planteamiento de numerosashipótesis sobre su mecanismo de acción en cerebro.Sin embargo, ha sido en la ultima década en donde, con laintroducción de nuevas técnicas en biologíacelular y molecular, se ha realizado un gran avance identificandonuevas dianas del etanol en cerebro. En este artículo se vaa revisar los nuevos hallazgos e hipótesis que permitenexplicar los diferentes efectos neurofarmacológicos deletanol, incluyendo aquellos involucrados en la tolerancia ydependencia y que pueden participar en el daño cerebral aque conlleva el consumo crónico de alcohol.
Mecanismos de laacción del etanol sobre el SNC
El etanol es unasustancia adictiva poco potente, si se considera que sus efectosfarmacológicos se observan tras la ingesta de gramos deetanol, mientras que en otras sustancias adictivas dichos efectosse observan con miligramos/peso corporal. A su vez, la complejidady multitud de efectos que induce se debe, en parte, a su estructuraquímica que le permite interaccionar con grupos polares y nopolares de componentes de la membrana, lo que explica su elevadasolubilidad tanto en un entorno acuoso como lipídico. Estapropiedad hace que el etanol, a diferencia de otras sustancias,atraviese libremente la barrera hematoencefálica y sedetecte en pocos minutos en cerebro tras la ingesta de alcohol. Lasinteracciones del etanol con los componentes de la membrana(especialmente con los lípidos), ha sido una barreraimportante para poder dilucidar los mecanismos de acción deletanol en cerebro, así como encontrar sus posiblesreceptores específicos. De hecho, se ha sugerido que eletanol es una sustancia sin «receptor». En losúltimos años, mediante la utilización deproteínas recombinantes, ha permitido abordar lainteracción del etanol con diferentes proteínas(canales iónicos, receptores, enzimas involucrados entraducción de señales) e investigar qué tipode receptores son diana del etanol1.
Todos estosdatos de los últimos veinte años, han conducido alplanteamiento de dos hipótesis sobre la acción deletanol sobre el SNC: 1) Interacción del etanol con loslípidos de membrana, 2) Acción del etanol sobrereceptores ligados a canales iónicos.
Interacción del etanol con los lípidos demembranas
Meyer y Overtonpostularon a principios de siglo que la potencia anestésicade algunos compuestos, incluyendo diferentes tipos de alcoholes, secorrelacionaba con el poder de solubilizar los lípidos demembrana. Esta correlación fue posteriormente corroboradapor Seeman en 19722, quien postuló que losanestésicos, además de modificar los lípidos,alteraban las proteínas de membrana. A partir de estemomento, un gran número de trabajos demuestran que eletanol, a altas concentraciones (intoxicación aguda), escapaz de fluidificar los lípidos y alterar lasproteínas de las membranas neuronales. Por el contrario, elconsumo crónico de alcohol conlleva a cambiosfísico-químicos de la membrana, induciendo una mayorrigidez en su estructura (tolerancia a la fluidificación deletanol) y afectando la conformación de proteínasembebidas en la bicapa lipídica. Se sugiere que estoscambios podrían participar en la tolerancia y dependencia aletanol3.
Trabajos de losúltimos diez años demuestran que estahipótesis tiene varias limitaciones: a) La primera se basaen que las concentraciones que se requieren para fluidificar loslípidos de membrana, son excesivamente elevadas (> 100mM), y a dosis relevantes farmacológicamente (< 50 mM),los efectos no son significativos4. b) Un aumento detemperatura corporal también induce fluidificación delos lípidos de la bicapa y no se observan cambios en elcomportamiento similares a los de la intoxicaciónalcohólica aguda. c) Datos recientes indican que losalcoholes de cadena corta (metanol, etanol) son potentesinhibidores de la función del receptor para glutamato,N-metil-D-aspartato (NMDA), mientras alcoholes de cadena larga, queposeen un elevado poder de solubilizar los lípidos demembrana, no inhiben la función de este receptor.
La idea queactualmente se tiene sobre la participación de estemecanismo, es que el etanol, a bajas concentraciones, podríainteraccionar con microdominios específicos de la membrana,modificando y modulando ciertas interaccionesproteína-membrana yproteína-ligando5,6.
Accióndel etanol sobre receptores asociados a canalesiónicos
Mediante lautilización de técnicas neurofisiológicas, enbiología celular y molecular, en los últimosaños se ha demostrado que el etanol a bajas concentraciones(10-25 mM), puede interaccionar y/o modular la función dealgunas proteínas de membrana, entre las que se incluyenreceptores para neurotransmisores asociados a canalesiónicos, así como ciertas proteínas queparticipan en procesos de transducción de señales1. La participación de cada uno de los sistemas oreceptores depende de la dosis/concentración, del tiempo deingesta de alcohol y del efecto farmacológico (ej.estimulación, acción ansiolítica,hipnótica, refuerzo).
Referente alugares específicos en los que se ha postulado que el etanolpodría interaccionar con receptores asociados a canalesiónicos serían (Fig. 1): A concentraciones elevadas(> 100 mM), el etanol interaccionaría con los restospolares de los fosfolípidos de membrana (mediante laformación de puentes de hidrógeno) modificandociertos microdominios de algunas proteínas. A bajasconcentraciones de etanol, con mayor relevancia a nivelfarmacológico (10-50 mM), la molécula de etanol puedeinteraccionar con los lugares hidrofóbicos localizados enlos dominios de las interfases agua-proteína oagua-lípidos (ej. los receptores GABAA yglicina), con los lugares cercanos al canal iónico (ej.receptor NMDA) o con el dominio extracelular alostéricoaminoterminal (ej. receptor para acetilcolina) (ver revisión6).
Figura 1. Representación esquemática de los posibleslugares de interacción del etanol con una proteínaasociada a un canal iónico. A altas concentraciones, eletanol puede interaccionar con los lípidos de la bicapalipídica1 y modificar microdominios de lainteracción lípido-proteína3, ointeraccionar con los grupos polares de losfosfolípidos2. A bajas concentraciones, se haobservado que el etanol puede interaccionar con la interfaselípido-agua4, alterando el mecanismo de aperturadel canal5. Se han observado tambiéninteracciones con lugares alostéricos de la proteínasituados en el dominio extracelular aminoterminal6(esquema modificado de la ref. 6).
Dentro de losreceptores asociados a canales iónicos, las proteínasque han recibido una particular atención y estudio durantelos últimos años han sido los receptores paraglutamato, NMDA y los receptores GABAA. Elinterés de la acción del etanol sobre estosreceptores reside en que el glutamato y el GABA son los principalessistemas de neurotransmisión en cerebro y alteraciones en lafunción y/o respuesta de estos receptores podríanexplicar ciertos efectos farmacológicos del etanol tanto enla intoxicación aguda como en los mecanismos de latolerancia y dependencia.
Etanol yreceptores NMDA
El glutamato esel principal neurotransmisor excitador en el SNC demamíferos y su acción se ejerce a través devarios receptores entre los que se encuentra el receptorN-metil-D-aspartato (NMDA). Este último receptor estáasociado a un canal de calcio y alrededor de éste selocalizan lugares de unión para el glutamato y otrosligandos (glicina, neuroesteroides, poliaminas) queactuarían modulando la acción del glutamato y laentrada de cationes en la célula (Fig. 2). La importancia deeste receptor reside en que la unión del glutamato alreceptor NMDA promueve la entrada de calcio en la célula,proceso que se ha demostrado ser crítico para lapotenciación a largo plazo, plasticidad sináptica yprocesos de memoria y aprendizaje7, y gran parte deestos procesos se ven gravemente afectados enalcohólicos.
Figura 2. Representación esquemática del receptorNMDA en la que se ilustra un canal central para cationes y loslugares para diferentes ligandos que modulan la acción delglutamato (esquema modificado de la ref. 8).
Evidenciasexperimentales de los últimos diez años handemostrado que tanto bajas como altas concentraciones de etanolinhiben la función del receptor NMDA. Los primeros estudiosque describieron este efecto se realizaron en células encultivo procedentes de hipocampo y cerebelo, observando queconcentraciones de etanol (5-10 mM) relevantes a nivelfisiológico inhibían la entrada de calcio en lacélulas, cuando se estimulaba el receptor NMDA. Estoshallazgos se confirmaron en diferentes preparaciones de cerebro derata, o «in vivo» tras la administraciónintraperitoneal de etanol, demostrando que el etanol es capaz debloquear la liberación de neurotransmisores dependiente dela estimulación de los receptores NMDA (ver revisiones 1, 6,8). Sin embargo, algunas evidencias posteriores sugieren que lasensibilidad de los receptores NMDA al efecto del etanol no eshomogénea en cerebro, ya que mientras en algunas regiones seobserva una marcada inhibición (ej. hipocampo,culículo inferior), en otras apenas se detecta dicho efecto(ej. septum lateral).
En losúltimos años el receptor NMDA se ha clonado ycaracterizado. Este receptor es una proteínaheterodimérica de varias subunidades, que a su vez poseendiferentes variantes. La subunidad NR1(compuesta por 8 variantesalternativas) parece forma complejos pentaméricos con lafamilia de subunidades NR2, de las que se conocen la NR2 A, B, C yD. La composición en subunidades (y en sus variantesalternativas) de los receptores NMDA varía dependiendo de lazona de cerebro, proponiéndose que la mayor o menorsusceptibilidad al etanol de los receptores NMDA estaríamediada por variaciones en la composición en subunidades desu estructura. De hecho, algunas evidencias indican que lacombinación de la subunidad NR1 con la variante NR2B, esparticularmente susceptible al efecto inhibitorio del etanol (verrevisión 8).
Todos estosdatos indican que los receptores NMDA son muy sensibles al etanol ysugieren su participación en los efectosfarmacológicos que se observan, tanto en laintoxicación aguda (altas dosis) como en el consumo moderadode alcohol (bajas dosis)1. De hecho, lapotenciación a largo plazo asociada a la activacióndel receptor NMDA está implicada en ciertos procesos dememoria y aprendizaje. Se ha demostrado que el etanol inhibe lapotenciación a largo plazo en neuronas de hipocampo tantoin vivo como in vitro. Por tanto, episodios depérdida de memoria transitoria que se observa en laintoxicación aguda podrían estar relacionados coninhibiciones puntuales del receptor NMDA enhipocampo3,6,8. Posiblemente, agentes como el Acamprosato, que parece facilita la función del receptorNMDA9, podría ser un tratamiento efectivo enestas situaciones.
Referente aotros receptores de glutamato, estudios recientes indican que losreceptores kainato también se inhiben por el etanol y que laactividad de la proteína quinasa C (PKC) así como losreceptores NMDA podrían modular el efecto deletanol.
Etanol yreceptores GABAA
Elácido *-aminobutírico oGABA es el principal neurotransmisor inhibidor en cerebro, y suactivación se produce a través de dos clases dereceptores el GABAA y el GABAB.
El receptorGABAA es una proteína oligomérica formadapor diferentes subunidades y en su estructura posee un canaltransmembrana permeable a iones cloro. Alrededor del canal seencuentran diferentes lugares de unión (Fig. 3) en los quese incluye el GABA, benzodiazepinas, barbitúricos yneuroesteroides10. La estimulación delGABAA da lugar a un incremento en la permeabilidad deiones cloro en la célula, causando unahiperpolarización de la membrana neuronal.
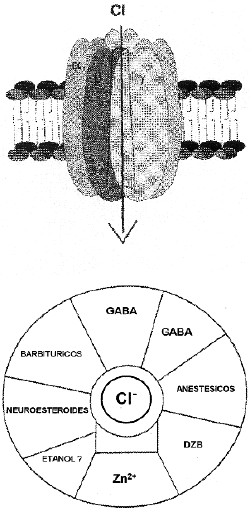
Figura 3. Esquema representativo de la estructura del receptorGABAA. En la parte superior se muestra cómo lasdiferentes subunidades se ensamblan alrededor del canal para ionescloro. En la parte inferior se representa los lugares paradiferentes ligandos que modulan la acción del GABA (esquemamodificado de la ref. 8).
Las similitudesde las acciones farmacológicas del etanol,barbitúricos y benzodiazepinas dieron lugar a las primerashipótesis sobre la acción del etanol en cerebro,sugiriendo que estas 3 sustancias compartían un mismomecanismo de acción sobre el receptor GABAA. Dehecho, desde la década de los 80 existenevidencias de que eletanol podría potenciar la acción GABAérgica.Estudios in vitro (en fracciones de cerebros aisladas océlulas en cultivo) e in vivo (mediante laadministración de 0,5 g de etanol/kg, a ratas) handemostrado que el etanol potencia el efecto del GABA sobre laactividad neuronal. Sin embargo, al igual que ocurre con el efectodel alcohol sobre los receptores NMDA, algunos estudios demuestrancierta variabilidad al efecto estimulante del etanol sobre elreceptor GABAA, dependiendo de la región decerebro estudiada. Estudios recientes indican que dichasvariaciones en la respuesta pueden estar relacionadas con lapresencia de determinadas subunidades del receptor GABAA(ver revisiones 1, 6, 8).
De hecho,mediante el uso de técnicas de DNA recombinante, se hademostrado que el receptor GABA posee múltiples subunidades(*, ß, *, *, y *) así como variossubtipos de cada subunidad (ej. *1 - *6), que in vivo se asocian en diversascombinaciones compuestas por subunidades y subtiposdiferentes10. La respuesta del receptor a diferentesefectores depende de la composición en subunidades delcomplejo proteico. Algunos estudios indican que el etanolinteracciona con un subtipo de receptor de GABAA, queposee la subunidad *6 y que solamente se expresa en célulasgranulares de cerebelo. Se ha propuesto que la presencia de estesubtipo en cerebelo podría estar implicado en lasalteraciones motoras que se observan en la intoxicaciónalcohólica aguda, ya que la administración deagonistas para benzodiazepinas las previenen. Otros hallazgosimportantes que apoyan la sensibilidad de alcohol por diferentessubunidades del receptor GABAA son los de Wafford y col.Estos autores expresaron las diferentes subunidades en oocitos deXenopus, demostrando que la acción del etanol sobreeste receptor requiere, de forma específica, la presencia dela isoforma *2L. Lasustitución de la isoforma *2L por *2S elimina el efecto del alcohol sobre elaumento en el flujo del ion cloro. Estas dos isoformas difierenentre sí por la presencia (*2L o isoforma larga) o ausencia (*2S o isoformacorta) de una inserción de 8 aminoácidos que poseenun consenso de fosforilación para la proteína quinasaC (PKC)11. Estudios recientes indican una menorsensibilidad a los efectos del alcohol, en ratones que carecen dela isoforma * de la PKC, y en lafunción del receptor GABAA en membranas aisladasde estos últimos animales12. Otros estudiosrecientes también indican que en neuronas de hipocampo elreceptor GABAB puede modular interacciones entre eletanol y el receptor GABAA.
En resumen, losdatos obtenidos hasta el momento indican que aunque lasinteracciones entre el etanol y el receptor GABAA soncomplejas, el etanol parece potenciar la acción del GABAsobre el receptor GABAA y que éste podríaser un mecanismo neurobiológico de la acciónansiolítica que produce el alcohol, ya que este receptorjuega un papel importante en la modulación de laansiedad.
Etanol yreceptores para serotonina
Desde hacedécadas se conoce que el sistema serotoninérgico encerebro desempeña un papel importante en ciertos efectosansiolíticos y de refuerzo asociados al consumo de alcohol.De hecho, un gran número de trabajos han demostrado unarelación entre los niveles de serotonina y consumo de etanolen animales experimentales y humanos13. Así,algunos animales con preferencia al etanol (animales P) poseen unosniveles de serotonina cerebrales aproximadamente un 20% másbajos que los animales con aversión por el alcohol (NP).Además, sustancias que inhiben la recaptación deserotonina (ej. fluoxatina, zimetidina) aumentan suconcentración en las terminales sinápticas ydisminuyen el consumo de etanol en roedores, pollos, monos yhumanos13.
Respecto alefecto del etanol in vitro sobre la función de losdiferentes receptores serotoninérgicos, concentracionesbajas de etanol no parecen afectar los subtipos de receptores5-HT1 y 5-HT2 en neuronas. Sin embargo,algunos trabajos sugieren que los receptores 5-HT2podrían estar implicados en el mecanismo de preferencia delconsumo de alcohol en roedores (ver revisiones 1, 6).
Estudiosrecientes indican que el etanol, a bajas concentraciones,interacciona con el subtipo 5-HT3, estimulando opotenciando su función. El receptor -HT3 es elúnico receptor de serotonina asociado a un canaliónico y se localiza principalmente en regiones del sistemamesolímbico de cerebro. Estudios electrofisiológicosrealizados en neuronas en cultivo y en células embrionariashumanas de riñón, transfectadas con un cDNA quecodifica la proteína del receptor 5-HT3,demuestran que concentraciones de 10-25 mM de etanol aumentan laentrada de iones a la célula, cuando son estimuladas conserotonina. Además, la presencia de un antagonista delreceptor 5-HT3 (ICS 205-930) elimina el efecto deletanol. Estos resultados indican que el etanol puede interaccionardirectamente con el receptor 5-HT3 y sugieren un posiblepapel de estos receptores en el consumo de alcohol. De hecho,resultados de Carboni y col. sugieren que existe unainteracción entre los sistemas serotonérgicos ydopaminérgicos y que ambos sistemas mediarían laspropiedades de refuerzo del etanol14. Estahipótesis la apoyan los resultados obtenidos tanto enanimales como en humanos, en donde la administración de unantagonista selectivo de los receptores 5-HT3, reduce elconsumo voluntario de alcohol15.
Accióndel etanol sobre los canales dependiente de voltaje
Se sabe que elcalcio es un ion de enorme importancia a nivel celular ya queregula numerosas funciones incluyendo liberación deneurotransmisores, activación de diferentes enzimasintracelulares, etc. Una de las vías en las que este ionentra en la célula es mediante canales dependientes devoltaje, de los que se han descrito varios tipos. Numerosostrabajos indican que altas concentraciones de etanol inhiben loscanales de calcio tipo L, N y T, pero no se observa efecto en losdel tipo P. Algunos autores sugieren que el etanol podríainteraccionar específicamente sobre la proteína delcanal iónico. Sin embargo, los mecanismos involucrados en laacción del etanol sobre la homeostasis del calcio y sutrascendencia en la intoxicación aguda, no estátotalmente esclarecida. Sin embargo, el consumo crónico dealcohol induce aumento en el número de estos canales,participando, junto con el aumento en receptores NMDA, en lahiperexcitabilidad que se produce en el alcoholismo crónico(ver revisión 16).
En losúltimos años se ha demostrado que el etanolpodría también interaccionar con determinado tipo decanales para potasio inhibiendo o activando, dependiendo delsubtipo de canal estudiado, el flujo de entrada de este ion a lacélula. Además, algunas evidencias indican que eletanol puede interaccionar con lugares específicos de laproteína de un canal de potasio, y si estos lugares semutan, se altera la sensibilidad del canal al efecto del etanol. Seha sugerido que diferencias alelicas en la composición yexpresión de diferentes canales de potasio podríanafectar la respuesta al etanol, y explicar las diferenciasindividuales y genéticas a los efectos del etanol (verrevisión 16).
Acciones deletanol sobre los sistemas dopaminérgico yopiáceo
Durante losúltimos años diferentes evidencias clínicas yexperimentales han demostrado que el sistema dopaminérgicomesolímbico está involucrado en la regulaciónde los mecanismos de recompensa y propiedades de refuerzo de muchassustancias adictivas, incluyendo el etanol. El sistemamesocorticolímbico, principal vía involucrada en elrefuerzo, proyecta sus terminaciones nerviosas desde el áreaventral (VTA) al núcleo accumbens, tubérculoolfatorio, cortex frontal y amígdala. Diferentes evidenciasexperimentales demuestran que la administración de bajasdosis de alcohol (0,25 g/kg), in vivo o intracranealmente,estimula la liberación de dopamina en el núcleoaccumbens17. Igualmente, la ingesta voluntaria dealcohol en animales con preferencia al etanol estimula laliberación de dopamina en el núcleo accumbens. Eletanol parece actuar en el área tegmental ventral,incrementando de forma indirecta la frecuencia de descarga de lasneuronas dopaminérgicas, mediante una atenuación deltono inhibitorio que ejercen las interneuronas GABAérgicas.Además, la liberación de dopamina inducida por eletanol parece está modulada por el sistema opioide. Laestimulación de los receptores opioides, localizados en elárea tegmental ventral, aumentan la frecuencia de descargade las neuronas dopaminérgicas. El etanol podríaaumentar la transmisión dopaminérgica, mediante unaestimulación del sistema opioide endógeno. De hecho,la administración de naltrexona, a dosis que bloquea losreceptores opiáceos µ y *, o de un antagonista delos receptores *, previene laliberación de dopamina en el núcleo accumbensinducida tras la administración de alcohol. Estos y otrosdatos apoyan la hipótesis de que los receptores opioidespodrían regular la liberación de la dopamina inducidapor el etanol en el núcleo accumbens, y que este mecanismopodría participar en las propiedades de refuerzo del etanol.Datos que apoyan esta hipótesis son los resultadosclínicos y experimentales que demuestran que laadministración de naltrexona reduce el consumo de alcohol(ver revisión 16). Otros autores postulan que lainteracción del etanol con los sistemas dopaminérgicoy serotonérgico serían las que median las propiedadesde refuerzo del etanol14.
Papel delacetaldehído en los efectos psicofarmacológicos deletanol
Desde hace 30años existen evidencias que apuntan que elacetaldehído (sustancia que procede del metabolismo deletanol) puede mediar algunos efectos psicofarmacológicos yneurotóxicos del etanol. Sin embargo, esta hipótesisha sido y sigue siendo controvertida, ya que en condiciones deconsumo de alcohol es difícil detectar acetaldehídoen cerebro. El acetaldehído que deriva del metabolismoperiférico del etanol penetra con dificultad la barrerahematoencefálica, puesto que posee una aldehídodeshidrogenasa que lo metaboliza, y se necesitan altasconcentraciones de acetaldehído en sangre (de 100-200µM) para que puedadetectarse en cerebro. Estas concentraciones no se llegan aalcanzar ni incluso tras el consumo de altas dosis de etanol. Sinembargo, se ha demostrado que cuando se incuban homogenizados decerebro de rata con etanol se produce acetaldehído, lo quesugiere que posiblemente el cerebro tenga sistemasenzimáticos que lo metabolicen18. De losdiferentes sistemas enzimáticos que participan en elmetabolismo del etanol, el alcohol deshidrogenasa tipo I (principalvía del metabolismo del etanol a nivel hepático), nose ha detectado en cerebro, el citocromo P4502E1 sí seencuentra en el sistema nervioso central e incluso se induce trasel consumo crónico de alcohol19, sin embargo suactividad es baja. Finalmente, el sistema catalasa no sólose expresa en cerebro, sino que su actividad es relativamenteelevada en este órgano. De hecho, existen evidencias queapoyan el papel de la catalasa como mediador en algunos efectosfarmacológicos y conductuales del etanol. Así,animales acatalasénicos o tratados con inhibidores de lacatalasa presentan alteraciones en la narcosis y en la actividadmotora inducidas por el etanol. Los autores que apoyan estahipótesis sugieren que posiblemente el acetaldehído,que se origine vía catalasa en cerebro o en determinadasregiones del sistema nervioso central, serían los quemediarían algunos efectos psicofarmacológicos yconductuales del etanol. Aunque esta hipótesis es atractiva,sin embargo se requiere más pruebas directas que demuestrenel papel de la catalasa cerebral y/o la acción delacetaldehído en algunos de los efectos conductuales deletanol (ver revisión 20).
Como resumena esta primera parte de la revisión podríamosconcluir, que aunque el alcohol es una sustancia sin receptorespecífico, evidencias de los últimos añosindican que el etanol a bajas (consumo moderado)1 oaltas concentraciones (intoxicación aguda)6interacciona, de forma específica, con ciertos dominios dereceptores asociados a canales iónicos, inhibiendo algunossistemas excitatorios (ej. NMDA) y activando a los inhibitorios(ej. GABAA). Además, el etanolsensibilizaría los sistemas opioide y serotonérgico,que a su vez activarían las neuronas dopaminérgicasdel sistema mesolímbico y que participarían en laspropiedades de refuerzo del etanol. De hecho, se hapostulado1 que los sistemas dopaminérgico yopioide podrían actuar como refuerzo positivo, sin embargolos receptores GABA y los receptores serotonérgicos parecenmodularían los niveles de ansiedad en animales y humanos, yse considera que median el refuerzo negativo en las acciones deletanol.
Laacción del etanol sobre algunas proteínas queparticipan en sistemas de transducción deseñales1 (adenilato ciclasa, PKC, PKA, etc.)así como el acetaldehído, tambiénpodrían participar y modular las acciones del etanol a niveldel sistema nervioso central.
Mecanismosinvolucrados en la tolerancia y dependencia al etanol
La tolerancia sedefine como la necesidad de aumentar la dosis o cantidad de alcoholo sustancia adictiva, para que se sigan manteniendo los mismosefectos. En este proceso interviene la toleranciafarmacocinética en la que se producen cambios en laabsorción, distribución y metabolismo de la sustanciao del etanol, y adaptaciones del sistema nervioso central o cambiosque se producen para compensar el efecto inicial de la sustanciaadictiva o del etanol (tolerancia celular).
La dependenciase podría definir como la aparición de signos ysíntomas de abstinencia cuando se suspende el consumo dealcohol o sustancia adictiva. Por tanto, el síndrome deabstinencia podría representar una inadaptación delsistema nervioso central a la eliminación del alcoholdespués de un período de adaptación durante eldesarrollo de la tolerancia. Sin embargo, se ha sugerido que latolerancia al etanol no siempre conlleva a la dependencia. En elcaso del alcohol se han descrito dos tipos de procesos en eldesarrollo de la tolerancia. Un proceso de habituación, queprogresivamente reduce la intensidad de la acción deletanol, y un proceso de adaptación homeostática en elque se observan efectos opuestos a la acción inicial deletanol. Ambos tipos de tolerancia pueden coexistir, aunque endeterminadas condiciones un tipo de tolerancia puede predominar ypuede o no estar asociado a la dependencia. Por ejemplo, laadministración de altas cantidades de alcohol durante uncorto período de tiempo, reduce la intensidad de laacción del etanol y es suficiente para inducir tolerancia enalgunos efectos conductuales del etanol (ej. alteraciones motoras,efecto hipnótico e hipotermia), pero no conlleva a ladependencia. La dependencia parece estar asociada con un proceso deadaptación homeostática en el que se producen cambiosneuroadaptativos en ciertos sistemas diana del etanol (ej.receptores MDA, GABAA, dopamina) que tratan de compensarla acción del etanol y se originan tras un consumo prologado(en humanos se necesitan varios años) de altas cantidades dealcohol.
Uno de losreceptores que se han asociado en el mecanismo de la tolerancia ydependencia al etanol son los receptores para glutamato, NMDA. Eletanol a bajas o altas concentraciones inhibe la función deeste receptor, sin embargo su consumo crónico induce unaumento en el número de estos receptores y este cambioparece estar relacionado con un incremento en diferentessubunidades de la proteína del receptor en algunasáreas de cerebro. En cultivos de neuronas de cerebelo se hademostrado que la exposición crónica al etanol, nosólo incrementa el número de receptores NMDA sinotambién su función, medida mediante el flujo decalcio intracelular. Estudios electrofisiológicos enhipocampo, demuestran que tras un consumo crónico dealcohol, el aumento de receptores NMDA junto con el de canales decalcio dependientes de voltaje (VSCC), ocasionan unahiperexcitabilidad en los componentes sinápticos, queestaría involucrada en la sintomatología delsíndrome de abstinencia al alcohol (ver revisiones 6, 8,21). Esta hipótesis la corroboran los datos que indican quela administración de un antagonista del receptor NMDAatenúan las convulsiones que se observan en elsíndrome de abstinencia en animales. Igualmente, laadministración de acamprosato (N-acetil homotaurinato decalcio) a animales experimentales parece mejorar lossíntomas de la abstinencia al etanol. Este compuesto pareceactuar sobre los receptores NMDA y los canales de calciodependiente de voltaje, modulando la entrada de calcio en lacélula y atenuando la hiperexcitabilidad del síndromede abstinencia al alcohol22. De hecho, resultadosclínicos y experimentales indican que el acamprosatoatenúa las propiedades de refuerzo del etanol disminuyendosu consumo en alcohólicos.
Adaptaciones ydisminuciones en la neurotransmisión GABAérgicaparece también participar en los mecanismos de tolerancia ydependencia al alcohol. Se ha sugerido que el consumocrónico de alcohol alteraría la respuesta delreceptor GABAA dando lugar a cambios en la sensibilidadde las acciones del alcohol sobre este receptor. De hecho, algunosresultados indican que el consumo crónico reduce larespuesta del receptor GABA a su agonista disminuyendo el flujo deiones Cl en preparaciones de cerebro. Sin embargo,otros datos indican que este tratamiento no altera ni elnúmero de receptores ni la afinidad de los ligandos a losreceptores GABAA en cerebro (ver revisión 21).Resultados recientes sugieren, que diferencias en lacomposición de subunidades del receptor GABAA enlas distintas partes de cerebro, así como la diferentesusceptibilidad al etanol por ciertas subunidades (ej. *1,*2L ),podrían explicar la discrepancia en los resultados. Dehecho, varios trabajos demuestran que el consumo crónico dealcohol reduce la expresión (proteína y RNAm) de lassubunidades *1 y*, mientras que incrementala ß y la *3 endeterminadas partes de cerebro. Cambios en la expresión deciertas subunidades inducidas tras el consumo de alcohol,podrían disminuir la respuesta del receptor GABAAy participar en los mecanismos de la tolerancia y dependencia aletanol. De hecho, la administración de sustancias queaumentan la transmisión GABAérgica, mejoran lossíntomas del síndrome de abstinenciaalcohólica, mientras que la administración decompuestos que inhiben la transmisión GABAérgicapotencian dichos síntomas (ver revisiones 6, 8,21).
Otro de lossistemas de neurotransmisión que sufre ciertos cambios o sehipersensibiliza durante el consumo prologado de alcohol, es elsistema implicado en los mecanismos de recompensa y refuerzo osistema dopaminérgico mesolímbico23. Comoya se ha comentado, el etanol estimula la liberación dedopamina en el núcleo accumbens, sin embargo tras un consumocrónico y posterior deprivación, se observa undecremento en la actividad de las neuronas dopaminérgicasdel área tegmental ventral (VTA). La disminución delos niveles de dopamina se ha relacionado con el aumento en eldeseo de beber durante el síndrome de abstinencia. Se hasugerido que una activación en los receptores NMDA en elnúcleo accumbens (NA) ejercería una accióninhibitoria sobre la liberación de dopamina en estaregión, proponiéndose que el aumento en lafunción y número de receptores NMDA (inducido por elconsumo crónico de alcohol) activaría lasinterneuronas GABAérgicas y éstas inhibiríanla liberación de dopamina en el NA durante la abstinencia aletanol21.
Resumiendo,numerosas evidencias sugieren que adaptaciones en el númerode receptores NMDA y canales de calcio, junto con unadisminución en la respuesta de los receptoresGABAA (en ciertas áreas de cerebro),podrían participar en los mecanismos de tolerancia ydependencia al alcohol. Durante la abstinencia al alcohol seproduciría un aumento de la neurotransmisiónexcitatoria con respecto a la inhibitoria, dando lugar a unahiperexcitabilidad del sistema nervioso central, tal y como lodemuestran
algunos síntomas del síndrome de abstinencia(convulsiones, hiperactividad, alteraciones cognitivas, etc.). Almismo tiempo, los mecanismos de refuerzo y deseo de reiniciar labebida, durante el síndrome de abstinencia, estaríamediado por una disminución en la actividad del sistemadopaminérgico mesolímbico, que a su vezestaría modulado por los receptores NMDA y sistema opioide.De hecho, resultados recientes indican que la naltrexona(antagonista no selectivo de los receptores opiáceos), esefectiva en el tratamiento de la dependencia alcohólica, yaque reduce el deseo de beber y el número de recaídas.Sin embargo, la limitada eficacia de este compuesto en algunosestudios sugiere que los receptores opiáceos noserían los únicos que participarían en losefectos de refuerzo del etanol, y que posiblemente otros sistemasde neurotransmisión podrían estar involucrados eneste mecanismo (glutamanérgico,serotonérgico).
Factoresgenéticos que afectan la sensibilidad al alcohol
En 1940,Jellinek ya propuso que el alcoholismo podría tener uncomponente genético y clasificó una categoríadenominada «alcoholismo familiar» que se caracterizabapor una edad de presentación temprana de alcoholismo y unaevolución particularmente grave. Estudios posterioresrealizados en Escandinavia con grupos de niños adoptados,hijos o no de alcohólicos, corroboraron que el alcoholismopresenta un componente genético y que es independiente delentorno familiar. Estudios más recientes, realizados por elgrupo de Schuckit demuestran que existen grupos de alto riesgo enlos que se observan una importante predisposicióngenética al alcoholismo. Estos trabajos indican que personasjóvenes con un historial de alcoholismo familiar son menossusceptibles a los efectos del alcohol (aumento desudoración, niveles hormonales, medidaselectrofisiológicas) que sus respectivos controles(jóvenes procedentes de familias que no poseen historial dealcoholismo familiar). Además, en un estudio de seguimientodurante diez años, este investigador demostró un altoporcentaje de alcohólicos en los individuos queposeían un componente familiar de alcoholismo y queinicialmente mostraban menor sensibilidad a los efectos delalcohol25.
La mayor o menorsensibilidad a los efectos del etanol y su influenciagenética se ha observado también en animales. Dehecho, actualmente existen cepas de ratas y ratones seleccionadasgenéticamente y que difieren en el mayor o menor consumo dealcohol de forma voluntaria, o en la mayor o menor sensibilidad alos efectos del etanol (ver revisión 26). Un dato que aportalos estudios en animales experimentales, es que lapredisposición genética a un efecto del etanol, noafecta necesariamente la respuesta a otros efectos. Estos estudiostambién nos sugieren que en las respuestas al etanol, como aotras sustancias adictivas, participan varios genes queinteractúan de forma compleja con la dotacióngenética de cada individuo y con el ambiente.
Sin embargo, enlos últimos años con la introducción deanimales knockout o animales en los que se les elimina un genconcreto, se ha podido estudiar la participación dediferentes genes en los efectos que produce el etanol. Así,se han encontrado alteraciones en la respuesta al etanol enanimales que carecen de los genes que codifican al: receptor5-HT1B, Fyn tirosina quinasa y a los receptores paradopamina D2, sugiriendo que estos genes modulan elcomportamiento inducido por el etanol en animales (verrevisión 26). Recientemente, Thiele y col.27 handescrito la participación del neuropéptido Y (NPY) enel consumo y efectos del etanol. El NPY está formado por 36aminoácidos y se ha demostrado que participa y modula muchosprocesos en los que se incluyen: estimula el apetito, facilita elaprendizaje y procesos de memoria, modula la locomoción,reduce la ansiedad, etc. Thiele y col. han demostrado que ratonesque carecen del NPY consumen mucho más alcohol, son menossensibles a los efectos hipnóticos de etanol, se recuperanantes de una intoxicación por alcohol, que los ratones queposeen el NPY. Además, estos autores demuestran que lasobreexpresión del NPY conlleva a efectos opuestos (consumenmenos alcohol, aumenta la sensibilidad a los efectoshipnóticos del etanol) a los que se observaban en ratonesdeficientes en este neuropéptido.
Actualmente conla aparición de nuevas técnicas en genética ybiología molecular, se está tratando de posicionar o«mapear» regiones cromosómicas que contienengenes que pueden estar involucrados a la diferente respuesta aletanol (y de otras sustancias adictivas) que se observan endeterminadas especies de animales. Mediante loci de caráctercuantitativo o QTL (quantitative trait loci), se estánempezando a identificar regiones de ciertos cromosomas y genes quepueden estar asociados con analgesia, consumo de alcohol, procesosde memoria y aprendizaje (ver revisión 28). Por tanto, esposible que en los próximos años se produzca un granavance en el conocimiento de los genes involucrados, tanto en losdiferentes efectos que produce el etanol como los que producenotras sustancias adictivas.
Consumo de alcohol ydaño cerebral
Larelación entre consumo de alcohol y alteracionesneurológicas se conoce desde finales del siglo pasado,aunque el mecanismo por los que el alcohol produce estas lesionestodavía se desconoce.
El consumo dealcohol induce degeneración neuronal y pérdida ensustancia blanca. La muerte neuronal así como cambios en elnúmero y disposición de las terminaciones nerviosases más marcada en ciertas áreas de cerebro, talescomo la corteza cerebral frontal, hipocampo y cerebelo (verrevisiones 6, 29). Interesantemente, estas áreas participanen procesos cognitivos, emocionales, de memoria y de movimiento,que son procesos que se ven gravemente afectados en elalcohólico. El patrón de consumo de alcohol, cantidady años de ingesta alcohólica así como factoresgenéticos, parecen contribuir en el establecimiento dediferentes lesiones neurológicas. Así, el consumo dealtas cantidades de alcohol, de forma ocasional, se ha asociado conconvulsiones, estados epilépticos y un aumento del riesgo dehemorragias cerebrales (ver revisión 30). Sin embargo,alteraciones importantes cognitivas y cerebrales sólo se hanobservado en individuos con una ingesta importante de variosaños de consumo. Mediante el uso de tomografía axialcomputerizada y tomografía de emisión de positrones,estudios recientes indican que el consumo crónico de alcoholdisminuye el volumen del cerebro y reduce el flujo y la tasametabólica cerebral. Algunos de estos efectos se reviertentras un período de abstinencia.
Referente a laposible relación entre consumo de alcohol y alteracionesneurológicas, en un estudio en donde se midieron 12variables diferentes de consumo de alcohol, no las pudieronrelacionar con atrofia cerebral. Sin embargo, estudios recientesencuentran una relación entre consumo de alcohol y cambiosen algunas áreas de cerebro particularmentedilatación del tercer ventrículo (ver revisión30).
Otras de laspreguntas que se ha planteado durante los últimosaños, es si el consumo moderado de alcohol induce o noalteraciones cognitivas o neurológicas. Sin embargo, losresultados publicados hasta el momento son controvertidos, ya quemientras algunos trabajos demuestran que el consumo de 30-60 g dealcohol/día se asocia con efectos negativos en lasestructuras cerebrales y en su función, otros trabajosindican que el consumo de 2 vasos de vino/día previene lademencia en individuos mayores de 65 años (verrevisión 1).
Todos estosdatos indican, que aunque el abuso prolongado de alcohol induceimportantes alteraciones cognitivas y neurológicas, noparece existir una clara relación entre cantidad de alcohol,años de consumo y alteraciones cerebrales, sugiriendo que,además del alcohol, existen otros factores de riesgo y/ogenéticos que pueden potenciar la acción del etanolen cerebro. Entre estos factores merecen destacar lossiguientes:
Factoresnutricionales. El etanol contiene calorías y su consumoconlleva a estados de desnutrición y carencia de vitaminas,algunas de ellas importantes para el funcionamiento del SNC.Así, por ejemplo, la deficiencia en tiamina que se observaen algunos individuos alcohólicos conlleva a laencefalopatía de Wernicke´s, que cursa con ataxia detronco, cambios oculares y confusión mental. Muchospacientes con esta encefalopatía desarrollan una psicosis deKorsakoff que se manifiesta por una grave amnesia anterograda omemoria a corto plazo, desorientación y polineuritis en losmiembros inferiores. Por tanto, factores nutricionales y dedeficiencias en vitaminas, tales como la tiamina, puede potenciarla acción del etanol en cerebro. Al mismo tiempo,deficiencias o anomalidades genéticas en la enzimatranscetolasa dependiente de tiamina, pueden jugar un papelimportante en la manifestación del síndrome deWernicke-Korsakoff en alcohólicos.
Factoresindividuales y genéticos. Tradicionalmente se dice quelas mujeres son más susceptibles al daño producidopor el consumo de alcohol, que los hombres. Esta diferenciaprincipalmente se ha establecido con la hepatopatíaalcohólica. Estudios recientes indican que las mujeres sontambién más susceptibles al daño cerebralinducido por el alcohol que los hombres. En dos estudios recientesse compararon las alteraciones neurológicas en hombres ymujeres alcohólicos, demostrando que con un consumosemejante entre ambos sexos las mujeres desarrollaban anteslesiones cerebrales que los hombres (ver revisión30).
Otra de lasalteraciones indirectas que produce el consumo de alcohol es laencefalopatía hepática, que se asocia condisfunciones hepáticas y posible acumulación deamonio a nivel cerebral. Aunque todavía desconocemos losmecanismos por los que ciertos individuos tienen ciertapredisposición a desarrollar cirrosis y/o alteracionesneurológicas, se piensa que ciertos factoresgenéticos podrían estar implicados en ambos tipos depatologías.
Mecanismosinvolucrados en la toxicidad del etanol en cerebro
Mediante el usode cultivos de diferentes tipos de células neuronales ygliales, en donde se pueden eliminar factores nutricionales,hormonales, genéticos, etc., se ha demostrado que el etanoles un tóxico celular y que, dependiendo de laconcentración, puede causar muerte o afectar laproliferación y diferenciación celular. Los estudiosen cultivos celulares también han permitido evidenciar quela sensibilidad de las células neurales al efectotóxico del etanol es mucho más marcada durante sudesarrollo, aunque la toxicidad también se manifiesta encélulas diferenciadas.
Referente a losmecanismos de toxicidad del etanol, se ha demostrado que tanto laintoxicación aguda como el consumo crónico de alcoholse produce un aumento en la generación de radicales librescon el consiguiente daño en las estructuras neuronales ygliales32. A su vez, el consumo crónico dealcohol causa un aumento en los receptores NMDA y canales deCa2+ dependiente de voltaje, que en ausencia de alcohol(síndrome de abstinencia al alcohol) ocasionarían unincremento en la entrada de calcio en la célula. Elevacionestransitorias del calcio intracelular desencadena una serie deprocesos (activación de radicales libres y proteasas,estipulación de segundos mensajeros, dañomitocondrial) que igualmente pueden conducir a la muerte neuronal.Un dato interesante, que apoyan algunos estudios recientes, es queel alcohol no sólo daña a las neuronas, sino que lascélulas gliales tienen una gran sensibilidad a los efectostóxicos del etanol31. De hecho, en un trabajoreciente33 se demuestra una importante reducciónde células gliales (37 %) en hipocampo de individuosalcohólicos, mientras que el número de neuronas enesta zona de cerebro no varió significativamente.Considerando que el hipocampo participa en funciones importantes,incluyendo procesos de memoria y aprendizaje y estas funciones seven gravemente afectadas en alcohólicos, los resultadossugieren que la reducción en células astroglialespodrían participar en estas patologías.
Agradecimientos
Algunos de lostrabajos que se refieren del grupo investigador que escribe estarevisión han sido financiados por las ayudas deinvestigación CICYT SAF96-0185, GV-D-VS-20-126-96 y DGESICPM97-0075.
Bibliografía
1. Eckardt MJ,File SE, Gessa GL, Hoffman PL, Guerri C, Kalant H, Koob GF, Li TK.,Tabakoff B. Effects of moderate alcohol consumption on the centralnervous system. Alcoholism: Clin Exp Res1988;22:998-1040.
2. Seeman P. Themembrane action of anesthetics and tranquilizers. Pharmacol Rev1972;23:583-655.
3. Guerri C,Grisolia S. Chronic ethanol treatment affects synaptosomalmembrane-bound enzymes. Pharmacol Biochem Behav1983;18:45-50.
4. Peoples RW,Li C, Weight FF. Lipid vs proin theories of alcohol action in thenervous system. Annu Rev Pharmacol Toxicol1996;36:185-201.
5. Barry JA,Gawrish K. Direct NMR evidence for ethanol binding to thelipid-water interface of phospholipid bilayer. Biochemistry 1994;33:8082-8.
6. Fadda F,Rossetti. Chronic ethanol consumption from neuroadaptation toneurogeneration. Prog Neurobiol 1998;56:385-431.
7. Sakimura K,Kutsuwada T, Ito I, Manabe T, Takayama C, Kushiya E, Yagi T, AizawaS, Inoue I, Suglyama H, Mishina M. Reduced hippocampal LTP andspatial learning in mice lacking NMDA receptor *1 subunit. Nature1995;373:151-5.
8. Faingold CL,Gouemo P, Riaz A. Ethanol and neurotransmitter interactions- frommolecular to integrative effects Prog Neurobiol 1998;55:509-35.
9. Madamba SG,Schweitzer P, Zieglgansberger W, Siggins GR. Acamprosate enhancesthe N-methyl-D-aspartate component of excitatory neurotransmissionin rat hippocampal Ca1 neurons in vitro. Alcohol. Clin ExpRes 1996;20:651-8.
10. MacdonaldRL, Olsen RW. GABAA receptor channels. Ann Rev Neurosci1994;17:569-602.
11. Wafford KA,Whiting PJ. Ethanol potentiation of GABA(A) receptors requiresphosphorylation of the alternatively spliced variant of the*2 subunit.FEBS Lett 1992;313:113-7.
12. Harris RA,McQuilkin SJ, Paylor R, Abeliovich A, Tonegawa S, Wehner JM. Mutantmice lacking the * isoform of protein kinaseC show decreased behavioral actions of ethanol and altered functionof *-aminobutyrate type Areceptors. Proc Natl Acad Sci USA 1995;92:3658-62.
13. Le MarquandD, Pihl RO, Benkelfat C. Serotonin and alcohol intake, abuse, anddependence: Findings of animal studies. Biol Psychiatry1994;36:395-421.
14. Carboni E,Acquas E, Frau R, Di Chiara G. Differential inhibitory effects of a5-HT3 antagonist on drug-induced stimulation ofdopamine-release. Eur J Pharmacol 1989;164:515-9.
15. Sellers EM,Tonestto T, Romach MK, Somer GR, Sobell LC, Sobell MB. Clinicalefficacy of the 5-HT3 antagonist ondansetron in alcoholabuse and dependence. Alcohol: Clin Exp Res1994;18:879-85.
16. Crews FT,Morrow AL, Criswell H, Breese G. Effects of ethanol on ionchannels. Int Rev Neurobiol 1996;39:283-367.
17. Imperato A,Di Chiara G. Preferential stimulation of dopamine release in thenucleus accumbens of freely moving rats by ethanol. J Pharmacol ExpTher 1986;239:219-28.
18. Aragon CMG,Rogan F, Amit Z. Ethanol metabolism in rat brain homogenates by acatalase-H202 system. Biochem Pharmacol 1992;44:93-8.
19. Montoliu C,Vallés S, Renau-Piqueras J, Guerri C. Ethanol-induced oxygenradical formation and lipid peroxidation in rat brain: effect ofchronic alcohol consumption. J Neurochem1994;63:1855-62.
20. Zimatkin SM,Deitrich RA. Ethanol metabolism in the brain. Addiction Biology1997;2:387-99.
21. Hoffman PL,Tabakoff B. Alcohol dependence: A commentary on mechanisms. Alcoholand Alcoholism 1996;31:333-40.
22. Spanagel R,Sieglgänsberger W. Anti-craving compounds for ethanol: newpharmacological tools to study addictive processes. TrendsPharmacol Sci 1997;18:54-9.
23. Di Chiara G,North RA. Neurobiology of opiate abuse. Trends Neurobiol Sci1992;13:185-93.
24. Goodwin,D.W. «Alcoholism and heredity». Arch Gen Psychiatr1979;36:57-61.
25. Schuckit MA.Low level of response to alcohol as a predictor of futurealcoholism. Am J Psychiatry 1994;151:184-9.
26. Crabbe JC,Phillips TJ. Genetics of alcohol and other abused drugs. Drug andAlcohol Dependence 1998;51:61-71.
27. Thiele TE,Marsh DJ, Marie LS, Bernstein IL, Palmiter RD. Ethanol consumptionand resistance are inversely related to neuropeptide Y levels.Nature 1998;396:366-9.
28. Crabbe JC,Phillips TJ, Buck KJ, Cunningham CL, Belknap JK. Identifying genesfor alcohol and drug sensitivity: recent progress and futuredirections. Trends Neuroscience 1999;22:173-9.
29. Harper C.The neuropathology of alcohol-specific brain damage, or doesalcohol damage the brain? J Neuropathol Ex Neurobiol 1998;57:101-10.
30. Neiman J.Alcohol as a risk factor for brain damage: neurologic aspects.Alcoholism: Clin Exp Res 1998;22:346S-51S.
31. Guerri C,Renau-Piqueras J. Alcohol, astroglia, and brain development. MolNeurobiol 1997;15:65-81.
32. Montoliu C,Vallés S, Renau-Piqueras J, Guerri C. Ethanol-induced oxygenradical formation and lipid peroxidation in rat brain: Effect ofchronic alcohol consumption. J Neurochem1994;63:1855-62.
33. Korbo L.Glial cell loss in the hippocampus of alcoholics. Alcoholism: ClinExp Res 1999;23:164-8.