






Intensification of agriculture and urban development are the main forces degrading natural ecosystems, particularly within species-rich countries. In this study, we determined the variation in bird communities between anthropogenic (i. e., cattle grazing lands, crop fields, urban areas) and riparian habitats within a highly humanmodified landscape in northwestern Colombia. For this purpose, we assessed differences in bird community diversity, structure, and composition among studied habitats. We also investigated similarities of surveyed habitats according to the habitat preferences of species. Results showed that riparian habitats play a fundamental role in human-dominated landscapes, as they maintain diverse and complex bird communities, support interior forest bird species, and might promote heterogeneous bird communities in nearby habitats. Cattle grazing lands also exhibited heterogeneous bird communities, which might be a result of the presence of tall trees, abundant shrub cover, and proximity of riparian habitats. Few species were over dominant within crop fields and urban areas due to a simple vegetation structure, leading to homogeneous avian communities where disturbed-site species thrive. Given that increasing human population depends on agricultural and urban ecosystems worldwide, there is an urgent need to foster management and conservation activities within such ecosystems to support wildlife and enhance human welfare.
La intensificación de la agricultura y el desarrollo urbano son los principales factores de degradación de los ecosistemas naturales, particularmente en los países ricos en especies. En este estudio determinamos la variación de las comunidades de aves entre sistemas antropogénicos (i. e., potreros, cultivos, áreas urbanas) y riparios en un paisaje altamente modificado por el ser humano al noroeste de Colombia. Para ello, evaluamos las diferencias en la diversidad, estructura y composición de las comunidades de aves de los sistemas estudiados. Asimismo, analizamos la similitud entre los sistemas de acuerdo a la preferencia de hábitat de las especies registradas. Nuestros resultados muestran que los sistemas riparios juegan un papel fundamental en el paisaje antropogénico, ya que mantienen comunidades de aves diversas y complejas, alojan especies asociadas al interior de bosques y podrían estar favoreciendo la prevalencia de comunidades de aves heterogéneas en los sistemas adyacentes. Los potreros también exhibieron comunidades de aves heterogéneas, lo cual pudo deberse a la presencia de árboles altos, una alta cobertura arbustiva y la proximidad de los sistemas riparios. Pocas especies fueron sobre-dominantes en los campos de cultivo y las áreas urbanas puesto que estos sistemas presentan una estructura de la vegetación simple, lo cual propicia que las comunidades de aves sean homogéneas y dominadas por especies asociadas a sitios perturbados. Dado que mundialmente la creciente población humana depende de los ecosistemas agrícolas y urbanos, es imperante desarrollar actividades de manejo y conservación asociadas a este tipo de sistemas, de tal manera que sea posible conservar la vida silvestre y mejorar la calidad de vida del ser humano.
The socioeconomic changes that occurred during the end of the past century drastically altered natural ecosystems at different scales, resulting in a worldwide reduction of biodiversity (Pimm et al., 1995). In particular, the intensification of agriculture and the exponential growth of urban development resulted in a high rate of land-use change, which is considered to be the main force degrading natural habitats (Foley et al., 2005; Hanski, 2005). This scenario has been particularly serious for species-rich countries, where negative impacts of agriculture and human population growth have been profound (Pimm et al., 1995; Engelman et al., 2000).
Research on human dominated landscapes has increased in the last decades (Karanth and DeFries, 2010). However, most studies focused on temperate regions and sought to understand the ecology of remnants of natural habitat (Stouffer and Bierregaard, 1995; Estrada et al., 1997; Beier et al., 2002). As a result, our understanding of human dominated landscapes where natural habitats have been drastically reduced or completely extirpated is limited. In order to enhance the biological value of such landscapes, it is critical to understand their associated ecological patterns and processes (MacGregor-Fors et al., 2009; Martin and Possingham, 2005). In particular, birds have been widely used as a study group within human dominated landscapes, as they quickly respond to habitat alteration at different scales (Furness and Greenwood 1993), are sensitive to human related disturbances (Fisher and Peterson, 1997), and exhibit specific habitat preferences for sites with distinct disturbance levels (Ortega-Álvarez and MacGregor-Fors, 2009).
In general, anthropogenic habitats that constitute human dominated landscapes exhibit less diverse and more dominated bird communities than natural habitats (McKay, 1980). Nevertheless, bird community responses to distinct anthropogenic habitats (e.g., crop fields, cattle grazing lands, urban settlements) are not completely understood, as current studies have reported contradictory results. For example, crop fields do not always exhibit more diverse and complex bird communities than cattle grazing lands as suggested by Morris (2000), as this pattern could vary in relation to the remaining vegetation structure and the location of cattle grazing lands across the landscape (Estrada et al., 1997; MacGregor-Fors and Schondube, 2011). Moreover, urbanization is not consistently more negative for birds than agricultural lands (McKay, 1980; Bellocq et al., 2008), as certain cultivation practices could be more detrimental for bird communities than urban development itself (Petit et al., 1999; Faggi et al., 2008). Finally, although riparian habitats have been highlighted as crucial for wildlife species (Naiman et al., 1993; Woinarski et al., 2000; Sabo et al., 2005), little is known about their importance for bird communities within extremely human-altered landscapes.
In this study, we determined variations in bird communities among anthropogenic and riparian habitats within a highly human-modified Neotropical landscape. For this, we assessed differences in bird community species richness, abundance, structure, and composition (both taxonomic and functional) among crop fields, cattle grazing lands, urban settlements, and riparian habitats in northwestern Colombia. Also, we investigated the similarities of surveyed habitats according to species habitat preferences. According to their vegetation structure, we predicted that riparian habitats would exhibit the richest, most even, and most complex bird communities among all surveyed habitats, followed by cattle grazing lands and crop fields. We expected urban settlements to exhibit the highest bird abundances due to thriving urban exploiter species within these particular habitats. Finally, we presumed that riparian habitats and urban settlements would exhibit unique bird communities according to species habitat preferences, as the former would be more often used by interior forest species, while generalist and disturbed-site species would heavily prefer the latter.
Materials and methodsStudy site. Colombia is one of the richest countries in the world for bird species. However, almost 32% of its surface has been transformed into agricultural land, thus posing a serious threat for wildlife (Etter, 1993). This research was carried out in the montane region of San Jerónimo, Cordoba State, in the Caribbean region of Colombia. The study site is characterized by a biota with Chocoanic-Amazonic-Magdalenic affinity (Hernández-Camacho et al., 1992). The original vegetation type of the region included tropical dry forests; however, this original landscape has been drastically altered since the middle of the last century, mainly for agricultural, logging, and through urbanization (Etter, 1993; Henao-Sarmiento et al., 2008). As a result, there has been an important decline in bird diversity in the region (Laurence and Bierregaard, 1997; Renjifo et al., 2000).
Avian surveys. We surveyed resident bird communities from June to July 2010 during peak bird activity (i. e., 06-10h), using 5-min unlimited-radius point-counts (sensuRalph et al., 1996; Huff et al., 2000). We were careful when defining the location of point-counts and when performing bird surveys in order to avoid registering species using different habitats from those that we were actively surveying. We focused our study on resident birds, as they are usually more negatively affected by anthropogenic disturbance than migratory species. Pointcounts were separated by a minimum distance of 250 m in order to assure data independence (Ralph et al., 1996). We surveyed point-counts only once in order to maximize the study area. At each point-count, a single experienced observer who was highly familiarized with local birds (MED-L) registered all seen and heard bird species that were actively using the habitat (e. g., perching, foraging, moving through the vegetation). Although point-counts have been widely used for counting birds (Ralph et al., 1996; Huff et al., 2000), they do not consider differences in detectability among species and habitats. Thus, their associated results might be useful, but should be interpreted with caution. Since tree and shrub vegetation components have been pinpointed as crucial for bird communities in anthropogenic habitats (Petit et al., 1999), we measured the following traits within a circle (25 m radius) at the center of each point-count as suggested by Ralph et al. (1996) to characterize surveyed habitats:1) maximum tree diameter at breast height (DBH); 2) minimum tree diameter at breast height; 3) tree cover; 4) shrub cover; 5) tree species richness; 6) shrub species richness; 7) tree density, and 8) maximum tree height.
To contrast bird communities between those characteristic habitats that comprised the focal human dominated landscape, we conducted bird surveys at anthropogenic (i. e., crop fields, cattle grazing lands, urban settlements) and riparian habitats (Fig. 1). Since forest remnants are scarce in the region and those left are occupied by illegal armed groups, we excluded these from our surveys. In order to account for a representative sample of bird communities, we conducted 30 independent point-counts at each habitat. Riparian habitats (∼40 ha) were characterized by the presence of native vegetation located along rivers, with the Pink Trumpet-tree (Tabebuia rosea) and the Kapoktree (Ceiba pentandra) as dominant species. Although human activities have altered the original structure of the vegetation, a complex vertical structure still defines these habitats. Cattle grazing lands (∼170 ha) retained large native trees (i. e., Royal Poinciana, Delonix regia, Yellow Mombin, Spondias mombin) and were usually covered by a complex herbaceous stratum grazed by cattle. Within crop fields (∼80 ha), low native plant cover persisted, as only cotton (Gossypium herbaceum) and corn(Zea mays) are cultivated throughout the region. Finally, urban areas (∼120 ha) were represented by heavily built sites with high levels of human activity. Surveys were specifically performed at green, residential, and commercial sites within the city of Monteria, which occupies a surface of 3 117km2 and contains approximately 409 500 people (Negrete-Barrera and Garces-Pretel, 2010). From their closest border, anthropogenic habitats were located at similar distances from riparian habitats (cattle grazing lands, 1.1km; crop fields, 1km: urban areas, 0.9km).

Data analysis. To compare bird species richness among surveyed habitats, we calculated average species richness (Sobs Mao Tau) and 95% confidence intervals using EstimateS (Colwell, 2005). To determine differences in bird abundance between habitats, we first transformed bird abundance values (log10) to fit a normal distribution and then performed an ANOVA. In addition, 95% confidence intervals were calculated for each average abundance value. Following Payton et al. (2003), we assessed statistical differences among calculated bird species richness and mean abundance values with a α<0.01 when confidence intervals did not overlap. In order to contrast bird community structures among habitats, we used rankabundance plots (Magurran, 2004). Then, we performed an ANCOVA to determine statistical differences among resultant slopes.
We compared the taxonomic composition of surveyed habitats by calculating the ecological distances of bird communities using Biodiversity R as an extension for R (Kindt and Coe, 2005; R Development Core Team, 2010). Ecological distances summarize the variation in species composition among communities by calculating a single distance statistic and displaying the resultant matrix in a dendrogram constructed by clustering methods (Kindt and Coe, 2005). In order to account for differences in the taxonomic composition of bird communities due to species abundances, we calculated ecological distances from both an abundance-based (Bray-Curtis ecological distance) and an incidence-based approach (Jaccard ecological distance). To determine functional similarity of bird communities, we compared bird abundances with different feeding preferences among habitats. For this, we classified each recorded species into trophic groups according to its primary feeding resource, which was determined bibliographically (Howell and Webb, 1995; Schulenberg et al., 2007; Hilty and Brown, 2009). Then, we performed a Bray-Curtis ecological distance analysis (Kindt and Coe, 2005) to compare the functional composition of surveyed land-uses.
We evaluated the similarity of bird communities in relation to species habitat preferences. For this we classified each recorded species in one of 5 different categories: 1) interior forest species (species associated with the core of original habitats); 2) edge forest species (species associated with original habitats but mainly abundant at their limits); 3) disturbed site species (species associated with altered habitats); 4) disturbed edge species (species associated with altered habitats but mainly abundant at their limits), and 5) generalist species (species present at both original and altered habitats). Categories used to classify species in relation to their habitat preferences were determined based on personal observations and the information provided by Hilty and Brown (2009), Howell and Webb (1995), and Schulenberg et al. (2007). Finally, we performed a Bray-Curtis ecological distance analysis to determine bird community similarity between habitats in relation to the abundance of species with different habitat preferences (Kindt and Coe, 2005).
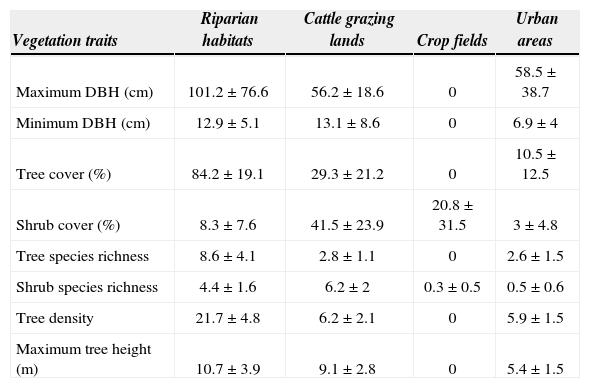
ResultsRiparian habitats showed a complex vegetation structure mainly related to the tree component, as wide, tall, and abundant trees of different species (Table 1) mostly covered these habitats. Although within cattle grazing lands trees were less abundant than in riparian habitats, medium-width tall trees were scattered amongst them. Moreover, cattle grazing lands contained an important shrub cover of different type of species. Crop fields were found to be very poor with regard to their vegetation traits, as only the shrub component was present and covered a small part of these habitats. Finally, urban areas contained a small number of short medium-width trees. In addition, the shrub component was widely undeveloped within urban areas.
Vegetation traits of surveyed habitats (average ± SD)
| Vegetation traits | Riparian habitats | Cattle grazing lands | Crop fields | Urban areas |
|---|---|---|---|---|
| Maximum DBH (cm) | 101.2±76.6 | 56.2±18.6 | 0 | 58.5±38.7 |
| Minimum DBH (cm) | 12.9±5.1 | 13.1±8.6 | 0 | 6.9±4 |
| Tree cover (%) | 84.2±19.1 | 29.3±21.2 | 0 | 10.5±12.5 |
| Shrub cover (%) | 8.3±7.6 | 41.5±23.9 | 20.8±31.5 | 3±4.8 |
| Tree species richness | 8.6±4.1 | 2.8±1.1 | 0 | 2.6±1.5 |
| Shrub species richness | 4.4±1.6 | 6.2±2 | 0.3±0.5 | 0.5±0.6 |
| Tree density | 21.7±4.8 | 6.2±2.1 | 0 | 5.9±1.5 |
| Maximum tree height (m) | 10.7±3.9 | 9.1±2.8 | 0 | 5.4±1.5 |
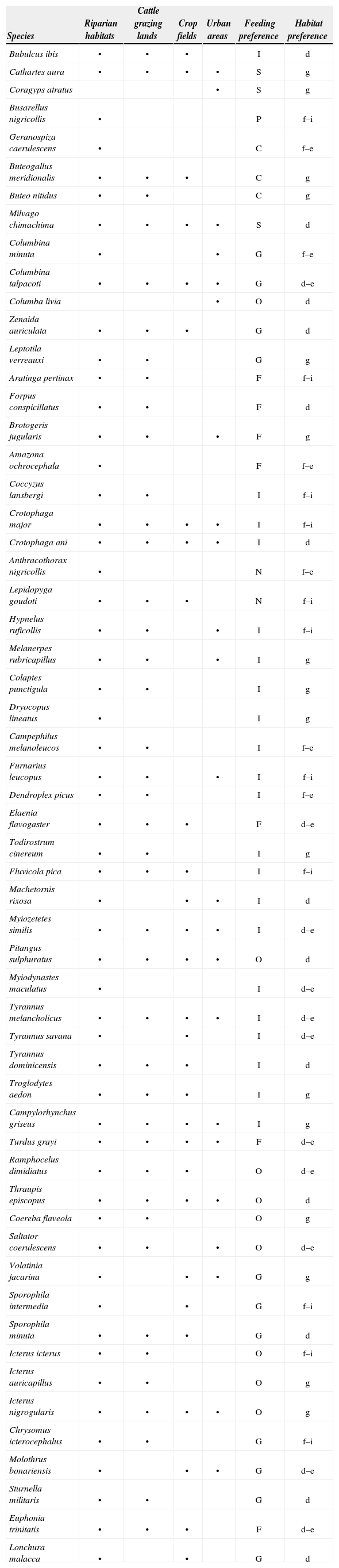
We recorded a total of 57 bird species among surveyed habitats (Table 2). According to the IUCN Red List of Threatened Species, all recorded species are classified under the “least concern” category (IUCN, 2011). Riparian habitats contained the highest number of bird species (55 spp.), followed by cattle grazing lands (42 spp.), crop fields (29 spp.), and urban areas (23 spp.). Recorded bird species were classified into 8 different groups according to their feeding preferences, with insectivores comprising the majority of recorded species (37%), followed by granivores (19%), omnivores (16%), frugivores (12%), carnivores (5%), scavengers (5%), nectarivores (4%), and piscivores (2%). In relation to species’ habitat preferences, 28% of the total number of recorded species were generalist species, 23% were disturbed site species, 19% were disturbed edge species, 19% were interior forest species, and 11% were edge forest species.
Bird species recorded in riparian habitats, cattle grazing lands, crop fields, and urban areas. Feeding and habitat preferences are given for each species. I= insectivore, G= granivore, O= omnivore, F= frugivore, C= carnivore, S= scavenger, N= nectarivore, P= piscivore, f-i= forest interior species, f-e= forest edge species, d= disturbed site species, d-e= disturbed edge species, g= generalist species
| Species | Riparian habitats | Cattle grazing lands | Crop fields | Urban areas | Feeding preference | Habitat preference |
|---|---|---|---|---|---|---|
| Bubulcus ibis | • | • | • | I | d | |
| Cathartes aura | • | • | • | • | S | g |
| Coragyps atratus | • | S | g | |||
| Busarellus nigricollis | • | P | f–i | |||
| Geranospiza caerulescens | • | C | f–e | |||
| Buteogallus meridionalis | • | • | • | C | g | |
| Buteo nitidus | • | • | C | g | ||
| Milvago chimachima | • | • | • | • | S | d |
| Columbina minuta | • | • | G | f–e | ||
| Columbina talpacoti | • | • | • | • | G | d–e |
| Columba livia | • | O | d | |||
| Zenaida auriculata | • | • | • | G | d | |
| Leptotila verreauxi | • | • | G | g | ||
| Aratinga pertinax | • | • | F | f–i | ||
| Forpus conspicillatus | • | • | F | d | ||
| Brotogeris jugularis | • | • | • | F | g | |
| Amazona ochrocephala | • | F | f–e | |||
| Coccyzus lansbergi | • | • | I | f–i | ||
| Crotophaga major | • | • | • | • | I | f–i |
| Crotophaga ani | • | • | • | • | I | d |
| Anthracothorax nigricollis | • | N | f–e | |||
| Lepidopyga goudoti | • | • | • | N | f–i | |
| Hypnelus ruficollis | • | • | • | I | f–i | |
| Melanerpes rubricapillus | • | • | • | I | g | |
| Colaptes punctigula | • | • | I | g | ||
| Dryocopus lineatus | • | I | g | |||
| Campephilus melanoleucos | • | • | I | f–e | ||
| Furnarius leucopus | • | • | • | I | f–i | |
| Dendroplex picus | • | • | I | f–e | ||
| Elaenia flavogaster | • | • | • | F | d–e | |
| Todirostrum cinereum | • | • | I | g | ||
| Fluvicola pica | • | • | • | I | f–i | |
| Machetornis rixosa | • | • | • | I | d | |
| Myiozetetes similis | • | • | • | • | I | d–e |
| Pitangus sulphuratus | • | • | • | • | O | d |
| Myiodynastes maculatus | • | I | d–e | |||
| Tyrannus melancholicus | • | • | • | • | I | d–e |
| Tyrannus savana | • | • | I | d–e | ||
| Tyrannus dominicensis | • | • | • | I | d | |
| Troglodytes aedon | • | • | • | I | g | |
| Campylorhynchus griseus | • | • | • | • | I | g |
| Turdus grayi | • | • | • | • | F | d–e |
| Ramphocelus dimidiatus | • | • | • | O | d–e | |
| Thraupis episcopus | • | • | • | • | O | d |
| Coereba flaveola | • | • | O | g | ||
| Saltator coerulescens | • | • | • | O | d–e | |
| Volatinia jacarina | • | • | • | G | g | |
| Sporophila intermedia | • | • | G | f–i | ||
| Sporophila minuta | • | • | • | G | d | |
| Icterus icterus | • | • | O | f–i | ||
| Icterus auricapillus | • | • | O | g | ||
| Icterus nigrogularis | • | • | • | • | O | g |
| Chrysomus icterocephalus | • | • | G | f–i | ||
| Molothrus bonariensis | • | • | • | G | d–e | |
| Sturnella militaris | • | • | G | d | ||
| Euphonia trinitatis | • | • | • | F | d–e | |
| Lonchura malacca | • | • | G | d |
Species richness, abundance, and the structure of bird communities. Rarefaction analysis revealed differences in bird species richness among surveyed habitats (Fig. 2a). Riparian habitats had the highest estimated bird species richness, followed by cattle grazing lands. Crop fields and urban areas did not differ in relation to their estimated number of bird species. Bird abundances also differed between habitats (F3, 114=12.3, p<0.001), being highest in crop fields, urban areas, and cattle grazing lands (Fig. 2b). Among all habitats, riparian habitats had the lowest bird abundance. The structure of bird communities and dominant bird species varied between surveyed habitats (F4, 144=99.5, p<0.001) (Fig. 3). The riparian bird community was the most even of all, followed by the community found in cattle grazing lands. Crop fields and urban areas bird communities exhibited a similar structure (Newman-Keuls: p=0.095), and were highly dominated by a few species.
 Estimated bird species richness for surveyed habitats. Comparisons of estimated bird species richness were performed using an abundance cut off point of n=198 (lesser total abundance recorded in riparian habitats). b) Bird abundances for surveyed habitats. Letters above values denote significant differences. Riparian= riparian habitats, grazing= cattle grazing lands, cropfield= crop fields, urban= urban areas.")
a) Estimated bird species richness for surveyed habitats. Comparisons of estimated bird species richness were performed using an abundance cut off point of n=198 (lesser total abundance recorded in riparian habitats). b) Bird abundances for surveyed habitats. Letters above values denote significant differences. Riparian= riparian habitats, grazing= cattle grazing lands, cropfield= crop fields, urban= urban areas.
 Tyrannus melancholicus and Leptotila verreauxi for riparian habitats; ii) Tyrannus melancholicus and Pitangus sulphuratus for cattle grazing lands; iii) Bubulcus ibis and Molothrus bonariensis for crop fields, and iv) Columba livia and Molothrus bonariensis for urban areas. Riparian= riparian habitats, grazing= cattle grazing lands, cropfield= crop fields, urban= urban areas.")
Rank-abundance plots of surveyed habitats. Dominant bird species for each surveyed habitat include: i) Tyrannus melancholicus and Leptotila verreauxi for riparian habitats; ii) Tyrannus melancholicus and Pitangus sulphuratus for cattle grazing lands; iii) Bubulcus ibis and Molothrus bonariensis for crop fields, and iv) Columba livia and Molothrus bonariensis for urban areas. Riparian= riparian habitats, grazing= cattle grazing lands, cropfield= crop fields, urban= urban areas.
Similarity of bird communities. Jaccard ecological distance showed that cattle grazing lands and riparian habitats contained very similar bird communities in relation to their taxonomic compositions (77% similarity), followed by crop fields (50% similarity), and by urban areas (36% similarity) (Fig. 4a). Bray-Curtis ecological distance also suggested that cattle grazing lands and riparian habitats shared taxonomically similar bird communities (69% similarity) (Fig. 4b). However, under this approach, crop fields and urban areas comprised a separate cluster (68% similarity), which was very different to that associated with cattle grazing lands and riparian habitats (46% similarity).
 Jaccard ecological distance dendrogram. b) Bray-Curtis ecological distance dendrogram. Urban = urban areas, Cropfield = crop fields, Riparian = riparian habitats, Grazing = cattle grazing lands.")
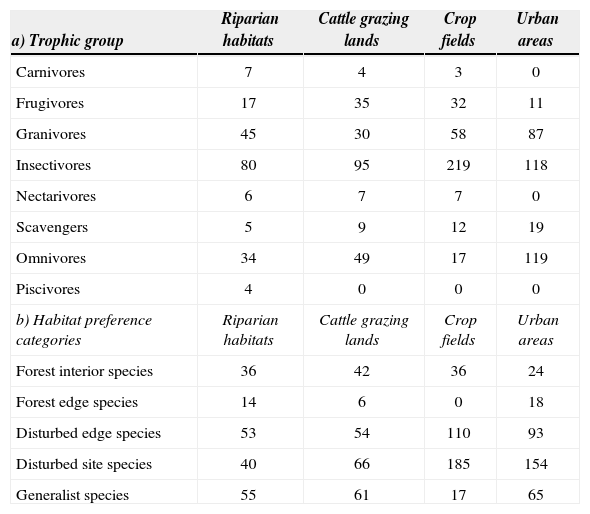
Riparian habitats and cattle grazing lands had the most functionally similar bird communities (83% similarity), followed by crop fields (68% similarity), and by urban areas (67% similarity). In particular, riparian habitats contained species from all trophic groups, with insectivores and granivores the most abundant (Table 3a). Cattle grazing lands were dominated by insectivore and omnivore species (Table 3a). With regard to crop fields, insectivores were widely dominant, followed by granivore species (Table 3a). Nectarivores, carnivores, and piscivores were absent from urban areas, while omnivores and insectivores dominated the community (Table 3a).
Abundance of a) trophic groups and b) species habitat preference categories among surveyed habitats. Reported values represent the sum of all recorded individuals for each group
| a) Trophic group | Riparian habitats | Cattle grazing lands | Crop fields | Urban areas |
|---|---|---|---|---|
| Carnivores | 7 | 4 | 3 | 0 |
| Frugivores | 17 | 35 | 32 | 11 |
| Granivores | 45 | 30 | 58 | 87 |
| Insectivores | 80 | 95 | 219 | 118 |
| Nectarivores | 6 | 7 | 7 | 0 |
| Scavengers | 5 | 9 | 12 | 19 |
| Omnivores | 34 | 49 | 17 | 119 |
| Piscivores | 4 | 0 | 0 | 0 |
| b) Habitat preference categories | Riparian habitats | Cattle grazing lands | Crop fields | Urban areas |
| Forest interior species | 36 | 42 | 36 | 24 |
| Forest edge species | 14 | 6 | 0 | 18 |
| Disturbed edge species | 53 | 54 | 110 | 93 |
| Disturbed site species | 40 | 66 | 185 | 154 |
| Generalist species | 55 | 61 | 17 | 65 |
According to the species’ habitat preferences analysis, surveyed habitats were separated into 2 different clusters that had a similarity value of 66%. The first cluster consisted of riparian habitats and cattle grazing lands (89% similarity), while the second was formed of crop fields and urban areas (83% similarity). Forest interior species were mainly recorded in riparian habitats and cattle grazing lands (Table 3b). Riparian habitats were highly used by generalist and disturbed edge species, while cattle grazing lands were dominated by disturbed sites and generalist species (Table 3b). Crop fields and urban areas were dominated by disturbed site species, followed by disturbed edge species (Table 3b).
DiscussionRiparian habitats were very important within the modified landscape, as they contained the highest estimated number of bird species among all surveyed habitats. Although riparian habitats have also been subjected to different negative anthropogenic impacts at the study site, they still contain a heterogeneous understory and canopy cover, which might enhance microhabitat segregation, benefiting a vast array of breeding bird species (Deschênes et al., 2003; Martin and Possingham, 2005). Cattle grazing lands also contained a high number of avian species, which may have been attracted from adjacent riparian habitats by their remaining tall trees and abundant shrub cover (Saab and Petit, 1992; Petit et al., 1999). As crop field vegetation was only composed of an herbaceous stratum dominated by a few crop species (i. e., Cotton, Corn), only a reduced number of bird species might be able to fulfill their habitat requirements within them (Petit et al., 1999; Benton et al., 2003; Berges et al., 2010). For urban areas, high levels of human related disturbances (e.g., pedestrians, cars), reduced vegetation cover, and the presence of exotic (i. e., Rock Pigeon – Columba livia) and brood parasite urban exploiter species (i. e., Shiny Cowbird – Molothrus bonariensis) could have caused the low species richness values for these habitats (Miller et al., 1998; Ortega-Álvarez and MacGregor-Fors, 2009).
Within crop fields, the availability of a limited number of feeding resources (i. e., grain, crop insects) could have promoted the over-dominance of particular gregarious species that are tolerant to human related disturbances (i. e., Shiny Cowbird, Cattle Egret - Bubulcus ibis) (Dhindsa and Saini, 1994). In the case of urban areas, urban exploiter species (i. e., Rock Pigeon, Shiny Cowbird) biased the increase of bird abundance values for the entire community (Ortega-Álvarez and MacGregor-Fors, 2009). These species thrive in urban systems because of a high availability of human related feeding resources, a decrease in competition with native species, and a reduction in predatory pressures (Shochat, 2004; Shochat et al., 2010). The more even bird community found in cattle grazing lands suggest that the presence of a greater variety of bird species contributed to the increase in bird abundances (Magurran, 2004). In this way, cattle grazing lands might offer a wider array of conditions and resources that are more evenly distributed among different bird species than within crop fields and urban areas. Finally, bird abundance values for riparian habitats were reduced, probably because of the complex and heterogeneous conditions and resources that occur within these habitats, which benefit no particular species and thus promote evenness in the community (Magurran, 2004).
Riparian habitats and cattle grazing lands had taxonomically similar bird communities. Proximity between cattle grazing lands and riparian habitats, as well as the presence of tall trees and a complex shrub stratum within cattle grazing lands, might facilitate the utilization of these 2 habitats by a similar number of taxonomically-related birds (Petit et al., 1999; MacGregor-Fors and Schondube, 2011). Incidence-based analysis showed that urban areas were taxonomically different from the rest of the surveyed habitats, which could be a result of the presence of urban exploiter species (i. e., Rock Pigeon, Black Vulture - Coragyps atratus) and the exclusion of a large number of native bird species (van Rensburg et al., 2009; MacGregor-Fors and Ortega-Álvarez, 2011). Nevertheless, the abundance-based analysis revealed a high similarity between urban areas and crop fields. This suggests that, although urban habitats and crop fields do not exactly contain the same type of bird species, they might promote high population numbers in some of those species shared (e. g., Shiny Cowbird, Smooth-billed Ani, Crotophaga ani; Cattle Tyrant, Machetornis rixosa; Ruddy Ground-dove, Columbina talpacoti). Such species could benefit from the reduction in competitive interactions with other species that are intolerant to the new conditions imposed by urban areas and crop fields (Khoury and Al-Shamlih, 2009; Shochat et al., 2010).
Insectivores were found to be very successful in all studied habitats. As human-modified ecosystems tend to contain large open areas, insectivore bird species with high visibility-dependent foraging strategies could have improved hunting success, and thus increased their population numbers (i. e., Tropical Kingbird, Tyrannus melancholicus, Great Kiskadee, Pitangus sulphuratus) (Saab and Petit, 1992; Hulme, 2007). Riparian habitats and cattle grazing lands were used by bird species with similar feeding preferences. This supports the idea that both habitats offer equivalent resources to birds as a result of their heterogeneous properties, which are mainly related to their vegetation structure (MacArthur and MacArthur, 1961; MacArthur et al., 1962; Martin et al., 2006). Although feeding resources can be found in large quantities within urban systems, only a small fraction of bird species can exploit them (e. g., omnivores, insectivores) (Shochat, 2004; Ortega-Álvarez and MacGregor-Fors, 2011a). Hence, it is not surprising that urban areas had the most dissimilar bird communities among all the surveyed habitats. As suggested by Dhindsa and Saini (1994), crop fields may offer concentrated amounts of food but only for particular groups of birds (i. e., granivores), which might result in the reduction of the functional heterogeneity of bird communities. However, our results suggest that crop fields still support bird communities that are functionally more heterogeneous than those present in urban areas.
Although landscape modification has reduced the abundance of interior forest species throughout the study site (Renjifo et al., 2000), some of these species were still recorded, mainly within riparian habitats and cattle grazing lands. The complex canopy cover of riparian habitats and the abundant understory cover present within cattle grazing lands may have been important for supporting interior forest species, as these offer different foraging, sheltering, and nesting sites (MacArthur et al., 1962; Saab and Petit, 1992; Scott et al., 2003). Riparian habitats and cattle grazing lands were also important for generalist species, which might have benefited from the presence of both the original and disturbed conditions that prevail in these habitats (Cerezo et al., 2009). High levels of human disturbance and the reduction of vegetation complexity, both characteristic of crop fields and urban areas, might have created conditions where the species that increase the most are the ones tolerant to continuous habitat disturbance.
Human dominated landscapes can still maintain high levels of biodiversity (Nowak and Nowak, 2006). As an increasing human population depends on these proliferating landscapes, there is an urgent need to understand related ecological patterns and processes in order to foster management and conservation activities focused on supporting wildlife and human welfare (Benton et al., 2003; Martin and Possingham, 2005; Berges et al., 2010). Although the short duration of our study might hamper the understanding of the annual variability of focal bird communities, our results represent a valuable instant frame that captures the shifts on bird community values among surveyed habitats, supporting and complementing those findings provided by previous studies from other regions of the globe. According to our results, riparian habitats play a fundamental role for avian communities in human dominated landscapes, as they maintain diverse and complex bird communities. Moreover, their protection and restoration is essential, as they may be important in supporting forest interior bird species and promoting heterogeneous bird communities in nearby habitats (Petit et al., 1999). Despite the fact that cattle grazing lands have been identified as detrimental for bird communities, the enhancement of tree and shrub cover could increase bird diversity values within these sites (Saab and Petit, 1992). To benefit birds, we recommend increasing the complexity of vegetation structure within crop fields, either by promoting multi-species crops or by incorporating live fences (Estrada et al., 1997). The promotion of vegetation complexity within urban areas, mainly related to the tree and shrub component, could favor more diverse bird communities by reducing suitable foraging habitats for urban exploiters (Ortega-Álvarez and MacGregor-Fors, 2011b), increasing the refuges against human disturbance for native birds (Fernández-Juricic et al., 2001), and promoting microhabitat diversification.
We are grateful to Susana I. Sevilla Beltrán, Bob Smith, and Yolanda Barrios for improving the English grammar of the manuscript.