






Se estudió la morfología y la anatomía foliar de los géneros del complejo Milla (Themidaceae) con el objetivo de identificar los caracteres foliares que permitan diferenciar entre los géneros y sus especies con énfasis en Milla biflora. Las muestras procedentes de campo y de herbario se procesaron a través de la microtecnia de inclusión en parafina. Las hojas son lineares, principalmente rollizas y subrollizas, excepto en Dandya balsensis, D. thadhowardii y Petronymphe que son triangulares y en algunas poblaciones de M. biflora son planas. El perfil de la lámina, visto en corte transversal, en general tiene forma depresa obovada, excepto en 3 especies, en que es obtriangular. El metaxilema en forma de “t” es un estado de carácter exclusivo de Bessera que permite diferenciarlo de los otros géneros que tienen metaxilema en forma “v”. No se encontraron caracteres exclusivos para el resto de los géneros del complejo. Con excepción de las especies D. hanniballi, M. mexicana y M. potosina y M. biflora, el mesofilo hueco es común en el resto de las especies en los diferentes géneros del complejo. La anatomía de la hoja permite reconocer, basándose en una combinación única de caracteres, a algunas especies de los diferentes géneros.
The aims of this study were to describe and compare foliar morphology and anatomy of the Milla complex genera (Themidaceae), in order to identify the foliar features that allow differentiating them. In addition, foliar variation was compared among genera and species with emphasis in Milla biflora. Field and herbarium samples were prepared with the paraffin microtechnique. Leaves are linear, mainly terete or subterete, except for Dandya balsensis, D. thadhowardii and Petronymphe with triangular shape and for some populations of M. biflora with flat shape. The lamina, seen in transverse section, has a depress-obovate shape, but 3 species have obtriangular. Metaxylem in a “t” shape is unique of Bessera and allows differentiating Bessera from the other genera with metaxylem in a “v” shape. Unique characters for the remainder genera of the complex were not found. With the exception of D. hanniballi, M. mexicana, M. potosina, and M. biflora mesophyll has a hollow in the other species of the Milla complex. Foliar anatomy allows to recognizing some species based make it a unique combination of characters.
Los géneros Bessera, Dandya, Milla, Jaimehintonia y Petronymphe conforman el complejo Milla y junto con el complejo Brodiaea constituyen a la familia Themidaceae (Fay y Chase, 1996; Pires et al., 2001; Pires y Sytsma, 2002). De los géneros del complejo en estudio, Milla es el que tiene un mayor número de especies, 10, algunas de ellas segregadas a partir de M. biflora Cav. (Gutiérrez y Solano, 2015; Howard, 1999; Ravenna, 1971), para Bessera se reconocen 3, para Dandya 4, y Jaimehintonia y Petronymphe son monotípicos. Los géneros del complejo Milla se caracterizan por ser hierbas geófitas con los cormos cubiertos por las bases membranosas de las hojas, las inflorescencias en forma de umbela con múltiples brácteas en la base y el ovario estipitado con el ginóforo adnado al tubo del perianto (Fay y Chase, 1996; Moore, 1953). Además, Moore diferenció los géneros del grupo por la forma del perianto, la longitud de los filamentos respecto al tamaño de la antera, la connación de los filamentos y el tamaño del ginóforo (Moore, 1953).
Algunos autores estudiaron a los representantes del complejo Milla para tratar de establecer sus relaciones filogenéticas (Fay y Chase, 1996; Gándara, Sosa y León de la Luz, 2009; Gándara, Specht y Sosa, 2014; Pires et al., 2001; Pires y Sytsma, 2002), sin que hasta el momento sean claras y de acuerdo con Gándara et al. (2014), los géneros no se recuperan. Un problema recurrente es la diferente interpretación en la morfología floral para algunos géneros del complejo (Gutiérrez, Terrazas, Hernández y Martínez-Cabrera, 2010). Por ejemplo, Lenz (1971) y Turner (1993) mencionan que existe connación de los estambres en los géneros del complejo Milla, pero solo se presenta dicha connación en las especies de Bessera (Gutiérrez et al., 2010). Ni la morfología ni la anatomía foliar se han evaluado con el fin de conocer su utilidad en la delimitación de los géneros y de las especies de este complejo.
La anatomía foliar proporciona caracteres en la sistemática de las plantas vasculares (Anderson y Creech, 1975; Gennaro, Pozner y Morrone, 2010; Goldblatt, Rudall y Henrich, 1990; Johnson, 1980; Kocsis, Darok y Borhidi, 2004; Mota-Alves, Scatena y Trovó, 2013; Rudall y Mathew, 1990; Rudall, 1991; Solano, Terrazas y González-Becerril, 2014). Esta fuente de datos apoya el reconocimiento de grupos a diferentes niveles taxonómicos en monocotiledóneas. Así, en Hyacinthaceae, Lynch, Rudall y Cutler, 2006 reconocen subfamilias, Tanker y Kurucu (1981) apoyan la delimitación de las secciones del género Allium, y otros autores (Brullo, Pavone y Spampinato, 1989; Ceja-Romero, Espejo-Serna y López-Ferrari, 2009; Echevarría, Morilla y Serrano, 1988; Membries, Pedrola-Monfort y Cuajapé-Castells, 2003) apoyan el reconocimiento de especies nuevas. Según lo anteriormente señalado, este estudio tuvo como objetivos describir la morfología y la anatomía foliar de los géneros del complejo Milla e identificar los caracteres foliar que permitan diferenciar entre los géneros y sus especies; así como evaluar la variación de M. biflora.
Materiales y métodosEn campo se recolectaron 19 especies de los 5 géneros del complejo Milla, poniendo énfasis en los representantes de M. biflora, especie de amplia distribución en la República Mexicana (Anexo). En cada localidad se recolectaron 3 individuos por especie, junto con los ejemplares de respaldo correspondientes que fueron depositados en los herbarios MEXU y FEZA (Holmgren, Holmgren y Barnett, 1990). Únicamente M. delicata H. E. Moore no fue localizada en campo, ni tampoco se pudo obtener material de ejemplares de herbario. Los individuos completos colectados en campo se fijaron en formalina-ácido acético-etanol (Ruzin, 1999). Una hoja de cada individuo se disectó, y de su tercio medio, del cual se separó un fragmento de 1cm aproximadamente, se deshidrató en un cambiador automático Leica (TP1020) con alcohol ter-butílico (20 a 100%) y se incluyó en parafina con punto de fusión de 56°C. Para 2 especies de B. tenuiflora (Greene) J. F. Macbr. y M. mortoniana H. E. Moore se extrajo una fracción de la lámina del tercio medio de un ejemplar de herbario (Anexo) y se proceso de forma similar a lo señalado anteriormente. Una vez incluidas las láminas foliares se realizaron cortes transversales de un grosor de 10-12μm con un microtomo rotatorio (Leica RM2125RT), se tiñeron con safranina y verde rápido (Ruzin, 1999) y se montaron en resina sintética. La morfología de la hoja se describió según Moore (1953). La forma del perfil de la hoja, visto en corte transversal, se adaptó de la propuesta de Radford, Dickinson, Massey y Bell (1974) para figuras planas (fig. 1), vistas en cortes transversales. Los tejidos de la lámina foliar se describieron de acuerdo con Lynch, Rudall y Cutler (2006).
. Los números indican la proporción longitud-anchura de la lámina.")
Forma geométrica de las hojas en sección transversal de los géneros y especies del complejo Milla. Tomadas y modificadas de Radford et al. (1974). Los números indican la proporción longitud-anchura de la lámina.
Se presenta una descripción morfológica y anatómica de la hoja para los géneros del complejo Milla. Los nombres de los géneros o especies se mencionan solamente en los casos donde existe variación intragenérica e interespecífica.
MorfologíaLas hojas son lineares, mayormente rollizas y subrollizas, en D. balsensis, D. thadhowardii y P. decora triangulares (fig. 2), y en M. biflora aplanadas (poblaciones 846, 902). La base de la lámina es aplanada e incolora, el ápice de la lámina es agudo en la mayoría de las especies, excepto en M. magnifica, M. mexicana y algunas poblaciones de B. elegans en las que es obtuso, el margen es entero, excepto en M. bryani, M. rosea, M. oaxacana y algunas poblaciones de M. biflora (896, 898, 988) en las que es involuto. D. balsensis, D. thadhowardii y P. decora presentan una quilla abaxial. La longitud de la lámina va de 50cm en M. filifolia y M. biflora (1388) hasta 100cm en M. magnifica. El diámetro de las hojas rollizas fluctúa entre 1mm en M. filifolia y 24mm en M. magnifica. Las especies con lámina triangular o aplanada tienen una anchura que va de 2-3mm en D. thadhowardii hasta 4mm en P. decora. El número de hojas por planta varía entre una en B. elegans y 7 en M. rosea. En general la lámina está postrada, y en ocasiones erecta antes de antesis o decumbente en algunas poblaciones de M. biflora (1156, 1257, 1344) (fig. 2). La lámina es glabra, con denticulaciones sobre los nervios excepto en algunas especies de Milla (M. filifolia, M. magnifica y una población de M. biflora 1196) donde la superficie es lisa.
 Bessera elegans (J. Gutiérrez et al., 1253) subteretes; B) Dandya thadhowardii (Gutiérrez 844) triangulares quilladas; C) Petronymphe decora (J. Gutiérrez y R. Ríos 1350) triangulares quilladas; D) Milla oaxacana (J. Gutiérrez et al., 1193) subrollizas; E) Milla biflora (Gutiérrez 1165) rollizas; F) Milla magnifica (J. Gutiérrez e I. Escalante 812) erectas subrollizas.")
Hojas del complejo Milla. A) Bessera elegans (J. Gutiérrez et al., 1253) subteretes; B) Dandya thadhowardii (Gutiérrez 844) triangulares quilladas; C) Petronymphe decora (J. Gutiérrez y R. Ríos 1350) triangulares quilladas; D) Milla oaxacana (J. Gutiérrez et al., 1193) subrollizas; E) Milla biflora (Gutiérrez 1165) rollizas; F) Milla magnifica (J. Gutiérrez e I. Escalante 812) erectas subrollizas.
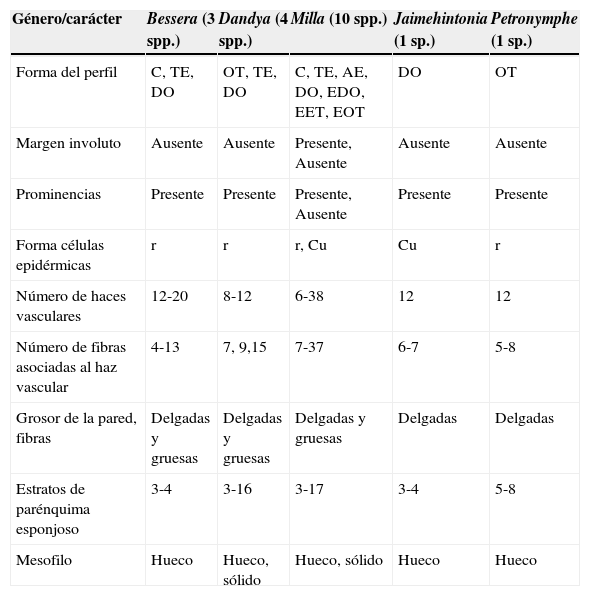
El perfil de la lámina en sección transversal varía (tablas 1 y 2; fig. 3A-D), y predomina la forma depresa obovada, seguida de la transversalmente elíptica y la circular. P. decora y 2 especies de Dandya se distinguen por su forma obtriangular (fig. 3B y 3E), mientras que M. biflora presenta todas las formas descritas para los otros géneros del complejo, excepto la obtriangular (tabla 2). La cutícula es estriada y delgada (< 1.5μm) en todos los materiales examinados. La epidermis es simple, y sus células vistas en sección transversal tienen forma cuadrada o rectangular con la pared anticlinal más larga (fig. 4A-C). En algunos puntos de la epidermis se observan 3 células que provocan un levantamiento debido a la elevación de la célula central que forma una protuberancia (figs. 3A, 3C, 3E, 4A y 4B) excepto en algunas especies de Milla (M. filifolia y M. magnifica) y algunas poblaciones de M. biflora en las que la epidermis es uniforme. Estas protuberancias en general se asocian a los haces vasculares y son conspicuas debido al alargamiento de la célula central (fig. 4A y 4B), la cual se clasifica en corta (< 50μm), larga (50-100μm) y muy larga (> 100μm). La pared periclinal externa de la epidermis es delgada o engrosada, las anticlinales generalmente delgadas, excepto en Bessera que tiene todas las paredes gruesas (fig. 4C). Cuando la pared es gruesa, esta siempre es de naturaleza celulósica. Los estomas se ubican en las células epidérmicas ordinarias, con prolongaciones anteriores y posteriores (fig. 4D), y vistos en superficie son paracíticos. El mesofilo es bifacial. El parénquima en empalizada es uniestratificado, con células uniformes excepto en D. balsensis, D. thadhowardii y P. decora, en que son más cortas en la cara abaxial. El parénquima esponjoso es compacto con células de menor a mayor tamaño hacia el centro de la hoja y de forma isodiamétrica. La parte más interna del mesofilo se colapsa o hay lisis celular y se forma un hueco (figs. 3A, 3B, 3C y 3F) excepto en 3 especies de Dandya (D. balsensis, D. hannibalii y D. thadhowardii), 3 especies de Milla (M. mexicana, M. mortoniana y M. potosina) y algunas poblaciones de M. biflora (fig. 3D y 3E). El mesofilo de todas las especies contiene rafidios dispersos (fig. 4E). Los haces vasculares son colaterales y se localizan subyacentes al parénquima en empalizada, se alternanen grandes y pequeños, y se disponen en una fila o formando un anillo en las hojas rollizas (fig. 3). El número de haces vasculares es variable dentro y entre géneros (tabla 1) y las especies de Milla muestran los extremos de la variación registrada (tabla 2). El metaxilema se dispone en forma de “t” en las especies de Bessera o de “v” en el resto de los taxa (fig. 4F y 4G) y el floema tiene tubos cribosos y células acompañantes muy pequeños comparados con los elementos del metaxilema (fig. 4H). El esclerénquima está conformado por paquetes de fibras que se ubican exclusivamente sobre el floema, con número y grosor variables. La mayoría de los materiales presentan pocas fibras (<15) con pared delgada o gruesa, Milla comúnmente tiene mayor abundancia de éstas (>20) y con paredes más gruesas (tablas 1 y 2; fig. 4I).
Características anatómicas de la hoja de los géneros del complejo Milla. Forma de la hoja en sección transversal.
| Género/carácter | Bessera (3 spp.) | Dandya (4 spp.) | Milla (10 spp.) | Jaimehintonia (1 sp.) | Petronymphe (1 sp.) |
|---|---|---|---|---|---|
| Forma del perfil | C, TE, DO | OT, TE, DO | C, TE, AE, DO, EDO, EET, EOT | DO | OT |
| Margen involuto | Ausente | Ausente | Presente, Ausente | Ausente | Ausente |
| Prominencias | Presente | Presente | Presente, Ausente | Presente | Presente |
| Forma células epidérmicas | r | r | r, Cu | Cu | r |
| Número de haces vasculares | 12-20 | 8-12 | 6-38 | 12 | 12 |
| Número de fibras asociadas al haz vascular | 4-13 | 7, 9,15 | 7-37 | 6-7 | 5-8 |
| Grosor de la pared, fibras | Delgadas y gruesas | Delgadas y gruesas | Delgadas y gruesas | Delgadas | Delgadas |
| Estratos de parénquima esponjoso | 3-4 | 3-16 | 3-17 | 3-4 | 5-8 |
| Mesofilo | Hueco | Hueco, sólido | Hueco, sólido | Hueco | Hueco |
AE: anchamente-elíptica; C: circular; Cu: cuadradas; DO: depresa-obovada; EDO: extensamente-depresa-obovada; EET: estrechamente-elíptica-transversalmente; EOT: estrechamente-oblonga-transversalmente; OT: obtriangular; r: rectangulares; TE: transversalmente-elíptica; TO: transversalmente-oblonga.
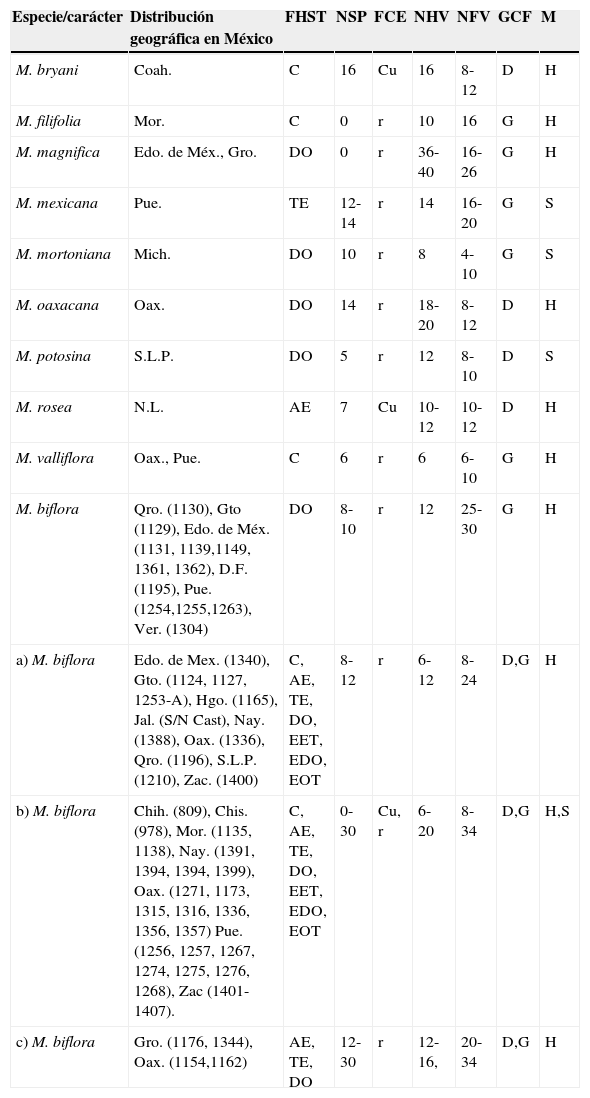
Características anatómicas de la hoja de 10 especies del género Milla.
| Especie/carácter | Distribución geográfica en México | FHST | NSP | FCE | NHV | NFV | GCF | M |
|---|---|---|---|---|---|---|---|---|
| M. bryani | Coah. | C | 16 | Cu | 16 | 8-12 | D | H |
| M. filifolia | Mor. | C | 0 | r | 10 | 16 | G | H |
| M. magnifica | Edo. de Méx., Gro. | DO | 0 | r | 36-40 | 16-26 | G | H |
| M. mexicana | Pue. | TE | 12-14 | r | 14 | 16-20 | G | S |
| M. mortoniana | Mich. | DO | 10 | r | 8 | 4-10 | G | S |
| M. oaxacana | Oax. | DO | 14 | r | 18-20 | 8-12 | D | H |
| M. potosina | S.L.P. | DO | 5 | r | 12 | 8-10 | D | S |
| M. rosea | N.L. | AE | 7 | Cu | 10-12 | 10-12 | D | H |
| M. valliflora | Oax., Pue. | C | 6 | r | 6 | 6-10 | G | H |
| M. biflora | Qro. (1130), Gto (1129), Edo. de Méx. (1131, 1139,1149, 1361, 1362), D.F. (1195), Pue. (1254,1255,1263), Ver. (1304) | DO | 8-10 | r | 12 | 25-30 | G | H |
| a) M. biflora | Edo. de Mex. (1340), Gto. (1124, 1127, 1253-A), Hgo. (1165), Jal. (S/N Cast), Nay. (1388), Oax. (1336), Qro. (1196), S.L.P. (1210), Zac. (1400) | C, AE, TE, DO, EET, EDO, EOT | 8-12 | r | 6-12 | 8-24 | D,G | H |
| b) M. biflora | Chih. (809), Chis. (978), Mor. (1135, 1138), Nay. (1391, 1394, 1394, 1399), Oax. (1271, 1173, 1315, 1316, 1336, 1356, 1357) Pue. (1256, 1257, 1267, 1274, 1275, 1276, 1268), Zac (1401-1407). | C, AE, TE, DO, EET, EDO, EOT | 0-30 | Cu, r | 6-20 | 8-34 | D,G | H,S |
| c) M. biflora | Gro. (1176, 1344), Oax. (1154,1162) | AE, TE, DO | 12-30 | r | 12-16, | 20-34 | D,G | H |
a) M. biflora=prominencias cortas (<50μm); b) M. biflora=prominencias largas (50-100μm); c) M. biflora=prominencias muy largas (>100μm).
AE: anchamente-elíptica; C: circular; Cu: cuadradas; D: delgada; DO: depresa-obovada; EDO: extensamente-depresa-obovada; EET: estrechamente-elíptica-transversalmente; EOT: estrechamente-oblonga-transversalmente; EPE: estratos de parénquima esponjoso; FCE: forma de las células epidérmicas; FHST: forma de la hoja en sección transversal; G: gruesa; GCF: grosor de la pared celular de la fibras; H: hueco; M: mesofilo; NFV: número de fibras asociadas al haz vascular; NHV: número haces vasculares; NSP: número de series de prominencias; r: rectangulares anticlinalmente; S: sólido; TE: transversalmente-elíptica.
 Bessera tuitensis (J. Gutiérrez et al., 1252) depreso-obovado; B) Petronymphe decora (Gutiérrez y R. Ríos 1350) obtriangular; C) Milla biflora (Castillejo s.n.) depreso-obovado; D) Milla biflora (J. Gutiérrez 1138) estrechamente-oblonga-transversalmente; E) Dandya thadhowardii (J. Gutiérrez 841) obtriangular; F) Milla biflora (J. Gutiérrez 1196) circular. Escala 300μm. es: esponjoso; flechas: protuberancias.")
Perfil de la lámina foliar de los géneros del complejo Milla, visto en sección transversal. A) Bessera tuitensis (J. Gutiérrez et al., 1252) depreso-obovado; B) Petronymphe decora (Gutiérrez y R. Ríos 1350) obtriangular; C) Milla biflora (Castillejo s.n.) depreso-obovado; D) Milla biflora (J. Gutiérrez 1138) estrechamente-oblonga-transversalmente; E) Dandya thadhowardii (J. Gutiérrez 841) obtriangular; F) Milla biflora (J. Gutiérrez 1196) circular. Escala 300μm. es: esponjoso; flechas: protuberancias.
 Bessera elegans (J. Gutiérrez y E. Bravo 1386) protuberancias epidérmicas (flecha); B) Milla biflora (J. Gutiérrez et al., 1162) protuberancias epidérmicas (flechas); C) B. elegans (J. Gutiérrez y E. Bravo 1386) paredes epidérmicas gruesas; D) Dandya balsensis (J. Gutiérrez 791) estomas con prolongaciones anteriores y posteriores; E), D. balsensis (J. Gutiérrez 791) rafidio (flecha); F) D. thadhowardii (J. Gutiérrez 798) haz vascular en forma de “v”; G) B. elegans (J. Gutiérrez y E. Bravo 1386) haz vascular en forma de “t”; H) D. hannibalii (J. Gutiérrez et al., 1241) floema con fibras en desarrollo; I) M. filifolia (J. Gutiérrez 1133) fibras de paredes gruesas sobre el floema. Escalas en A, B=100μm; F, G=50μm; C, E, H, I=20μm; D=10μm. e: epidermis; es: esponjoso; f: fibra; fl: floema; lpx: laguna del protoxilema; mx: metaxilema.")
Detalles anatómicos de la lámina foliar de los géneros del complejo Milla, vistos en sección transversal. A) Bessera elegans (J. Gutiérrez y E. Bravo 1386) protuberancias epidérmicas (flecha); B) Milla biflora (J. Gutiérrez et al., 1162) protuberancias epidérmicas (flechas); C) B. elegans (J. Gutiérrez y E. Bravo 1386) paredes epidérmicas gruesas; D) Dandya balsensis (J. Gutiérrez 791) estomas con prolongaciones anteriores y posteriores; E), D. balsensis (J. Gutiérrez 791) rafidio (flecha); F) D. thadhowardii (J. Gutiérrez 798) haz vascular en forma de “v”; G) B. elegans (J. Gutiérrez y E. Bravo 1386) haz vascular en forma de “t”; H) D. hannibalii (J. Gutiérrez et al., 1241) floema con fibras en desarrollo; I) M. filifolia (J. Gutiérrez 1133) fibras de paredes gruesas sobre el floema. Escalas en A, B=100μm; F, G=50μm; C, E, H, I=20μm; D=10μm. e: epidermis; es: esponjoso; f: fibra; fl: floema; lpx: laguna del protoxilema; mx: metaxilema.
Con esta investigación se confirma que las hojas en los géneros del complejo Milla son lineares, pero varían en la forma tridimensional: rolliza, subrolliza, plana o triangular. Éstas 2 últimas se describieron previamente (Howard, 1999; Moore, 1953). Las formas de la lámina señaladas son comunes en diversos géneros de Asparagales (Blunden, Yi y Jewers, 1973; Lynch et al., 2006; Sandoval-Zapotitla, García-Cruz, Terrazas y Villaseñor, 2010; Tanker y Kurucu, 1981). Así, en Hyacinthaceae las hojas son rollizas, subrollizas y aplanadas. En Agavaceae y Orchidaceae con frecuencia son aplanadas, mientras que en Iridaceae son aplanadas, cuadradas, plegadas, conduplicadas o rollizas (Rudall, 1991) y en Alliaceae, familia donde previamente se había clasificado a Milla, son aplanadas, rollizas y subrollizas (Dahlgren, Clifford y Yeo, 1985). De las 4 formas de la lámina descritas para los géneros del complejo Milla, las hojas rollizas y subrollizas muestran amplia variación; no obstante, mediante la forma del perfil, visto en corte transversal, se distinguen grupos bien definidos. Así, las hojas rollizas se clasifican como circulares (M. filifolia) o anchamente elípticas (M. rosea), mientras que las subteretes se clasifican como extensamente-depresa-obovada (poblaciones de M. biflora S/N Castillejos, 1388, 1400, 1405), transversalmente-elíptica (B. elegans 1386, 1252, D. hannibalii, M. biflora 1176, 1257, 1399, 1407 y M. mexicana), depresa-obovada (B. elegans 802, B. tuitensis, D. purpusii, Jaimehintonia, M. magnifica, M. oaxacana, M. potosina y poblaciones de M. biflora 1129, 1130, 1165, 1391, 1400, 1254, 1263) y estrechamente-elíptica-transversalmente (poblaciones de M. biflora 1240, 1253,1257, 1357). En este sentido la separación en diferentes estados de carácter del perfil de la hoja ofrece información adicional importante.
En los géneros del complejo Milla la epidermis es simple, similar a la de otros géneros de Asparagales (Lynch et al., 2006; Rudall, 1991); además, es habitual la presencia de protuberancias en la superficie de la epidermis, que se registran solamente en algunas especies de Tigridieae (Rudall, 1991) y Paepalanthus (Mota-Alves et al., 2013). Es de suma importancia identificar estas protuberancias en cortes transversales, ya que vistas en superficie se han descrito como denticulaciones hialinas con ayuda del microscopio estereoscópico (Moore, 1953) y pueden llegar a confundirse con papilas. Por su longitud, fue posible clasificar las protuberancias epidérmicas en 3 grupos que permiten separar a las poblaciones de M. biflora; además, las protuberancias cortas son el estado más común en el complejo Milla. La presencia de pared primaria gruesa, de naturaleza celulósica en la epidermis se interpreta como colenquimatosa y es habitual en las especies del complejo. Esta epidermis colenquimatosa probablemente le confiere a la lámina rigidez y flexibilidad; por ejemplo, las especies de Bessera tienen todas las paredes de su epidermis colenquimatosas y hacen que las hojas sean erectas antes de la antesis.
El desarrollo de un estrato de parénquima en empalizada es común en todos los taxa; mientras que el tamaño de sus células es homogéneo en la mayoría de los géneros, excepto en D. balsensis, D. thadhowardii y P. decora que presentan células del empalizada más cortas en la cara abaxial. Es possible que el cambio de tamaño se relacione con la forma obtriangular de la lámina. En los géneros del complejo Milla, la condición más común es que el parénquima esponjoso se colapse y forme un hueco, lo cual coincide con lo descrito para los géneros Allium, Asphodelus y Sisyrinchium (Goldblatt et al., 1990; Tanker y Kurucu, 1981). El colapso del parénquima esponjoso y la lisis celular son característicos de las hojas fistulosas (fig. 3A-C y 3F). Este carácter también se presenta en hojas xeromórficas como en Eriocaulon fistulosum (Eriocaulaceae), Juncus spp. (Juncaceae) y Gagea bohemica (Liliaceae) (Niklas, 1991; Peruzzi, Peterson, Tison y Peterson, 2008). La presencia de hojas fistulosas en el complejo Milla posiblemente sea una adaptación a ecosistemas secos para evitar la pérdida de agua, mantener una menor temperatura respecto al ambiente y contener mayor CO2 para la fotosíntesis (Gibson, 1996). En los miembros del complejo Milla, la presencia de hojas bifaciales se correlaciona con la condición fistulada. La correlación de estos caracteres posiblemente sea una novedad evolutiva para los géneros del complejo, pero para confirmar esta hipótesis es necesario evaluar la anatomía de los géneros del complejo Brodiaea tanto con hojas aplanadas como subrollizas (Hoover, 1939).
Los haces vasculares colaterales presentes en los géneros del complejo Milla tienen, con excepción de Bessera, el metaxilema dispuesto en forma de “v” (fig. 4F), distribución común en algunas Iridaceae (Goldblatt et al., 1990), donde se denomina “en forma de arco”. Por otra parte, Rudall (1991) señala que la posición de las fibras en el haz vascular tiene importancia taxonómica para distinguir especies, y se ha utilizado para Sisyrinchium (Ceja-Romero et al., 2009). Sin embargo, en las especies del complejo Milla, las fibras siempre se sitúan sobre el floema y en general son escasas; se ha observado que la presencia de abundantes fibras con paredes gruesas se relaciona con el desarrollo de hojas grandes, lo que favorece un mayor soporte. Lo anterior se presenta generalmente en los taxa que desarrollan hojas de más de 50cm de longitud como en B. elegans (80cm), M. biflora (75cm), M. magnifica (100cm) y M. mexicana (80cm).
Diferencias entre géneros y especiesBessera es el único género que presenta metaxilema en forma de “t” (fig. 4G) y todas las paredes de la epidermis colenquimatosas. Los otros géneros carecen de caracteres exclusivos; sin embargo, hay especies que pueden reconocerse por la combinación única de atributos foliares. Por ejemplo, D. hannibalii y D. purpusii, vistos en sección transversal, tienen el perfil de la lamina transversalmente elíptico, al igual que Bessera y Milla. En cambio, D. balsensis y D. thadhowardii presentan el perfil de la lámina obtriangular, semejante a Petronymphe, así como abundante tejido parenquimatoso dispuesto a lo largo de la costa (fig. 3B). Jaimehintonia gypsophila, B. tuitensis, y algunas poblaciones de B. elegans y de M. biflora (1129, 1130, 1165, 1391, 1400, 1254, 1263) comparten el perfil de la lámina depresa obovado (tabla 2). Los haces vasculares son por lo general más numerosos en Bessera (12-20) y Milla (6-38), y Petronymphe y Jaimehintonia comparten la presencia de 12 haces vasculares, mientras que Dandya tiene de 8-12 haces. Bessera, Dandya y Milla presentan fibras con paredes más gruesas y en mayor número que Jaimehintonia y Petronymphe y el mesofilo sólido agrupa a 3 especies de Dandya (D. balsensis, D. hannibalii y D. thadhowardii), M. mexicana, M. potosina y 2 poblaciones de M. biflora (1127 y 1271).
Diferencias interespecíficas en MillaEn las especies del género Milla, la anatomía foliar revela que existe una combinación única de caracteres que permite reconocerlas (tabla 2), excepto M. biflora. Esta especie, de amplia distribución geográfica, desde el sur de EUA hasta Honduras, tiene todos los estados de carácter anatómicos observables en el resto de las especies. Moore (1953) describió M. biflora como una especie polimórfica, con hojas estrechamente lineares, canaliculadas en la cara superior y redondeadas en la inferior, con diminutas denticulaciones hialinas en los nervios o algunas veces ausentes. Esta variación morfológica se confirma aquí y, además, se reconocen otras formas (tabla 2). Gándara et al. (2014) sugieren que M. biflora no presenta la misma historia evolutiva. Los resultados aquí presentados apoyan esta aseveración, ya que la anatomía foliar es variable en las poblaciones de M. biflora. Con base en la variación foliar de esta especie se reconocen 4 grupos, será de suma importancia poner a prueba estos caracteres foliares en un análisis de parsimonia.
Como ya se mencionó, las prominencias epidérmicas cortas es el estado de carácter más común en el complejo (B. elegans, D. balsensis, D. thadhowardii, Jaimehintonia, Petronymphe, M. potosina, M. rosea y poblaciones de M. biflora [“A”]). Las prominencias muy largas solo se presentan en 4 poblaciones de M. biflora (“C”) y las prominencias largas (50-100μm) las comparten las poblaciones M. biflora que se distribuyen en la Faja Volcánica Transmexicana y el grupo “B” de M. biflora (tabla 2).
Howard (1999) describió M. filifolia, M. mexicana y a partir de segregaciones de poblaciones de M. biflora, basándose en caracteres florales y vegetativos. Este autor señaló que en M. filifolia las hojas son subteretes diminutamente acanaladas adaxialmente y convexas abaxialmente; en M. mexicana, teretes o subteretes, y en M. potosina, cóncavo-convexas. La anatomía foliar de estas especies revela que el perfil de la lámina es transversalmente elíptico en M. mexicana y depresa obovada en M. potosina (tabla 2). M. filifolia exhibe las hojas más delgadas (≤1mm), de forma circular en sección transversal y solo comparte con M. magnifica la epidermis sin protuberancias.
Milla magnifica es la especie de mayor porte con hojas de mayor longitud (100cm) y un número de haces vasculares importante (36-40), habita en el bosque tropical caducifolio y se desarrolla en suelos ricos en materia orgánica, condiciones ecológicas que seguramente contribuyen a determinar su morfología. M. mexicana también se desarrolla en bosque tropical caducifolio; no obstante, difiere de M. magnifica en el tamaño de sus hojas (80cm), el perfil transversalmente elíptico, la epidermis con protuberancias, el menor número de haces vasculares (14) y el mesofilo compacto (tabla 2). El mesofilo compacto y las células epidérmicas rectangulares son caracteres distintivos en M. mexicana y M. potosina; sin embargo, esta última presenta un menor número de protuberancias epidérmicas (5), haces vasculares (12), fibras asociadas al haz vascular (8-10) y fibras con paredes delgadas.
De acuerdo con Moore (1953), M. bryani y M. rosea comparten la presencia de pedicelo y un número reducido de flores, así como su distribución geográfica. La anatomía foliar reveló que también comparten la forma del perfil y el margen involuto en la cara adaxial; pero M. bryani presenta un mayor número de series de prominencias (16 vs. 7) y haces vasculares (16 vs. 10-12). Ravenna (1971) describió M. oaxacana, basándose en diferencias morfológicas florales de poblaciones que Moore (1953) reconoció como parte de M. biflora. Aquí se muestra que M. oaxacana difiere de M. biflora por la presencia de 14 series de prominencias epidérmicas y el margen involuto en la cara abaxial (tabla 2). La morfología y la anatomía foliar proporcionan caracteres adicionales útiles para distinguir entre algunas especies del complejo Milla mediante una combinación única de caracteres. En cuanto al género, únicamente Bessera tiene atributos foliares exclusivos.
Esta contribución es parte de los requisitos del primer autor para obtener el grado de Doctor en Ciencias en el Programa del Posgrado en Ciencias Biológicas (PCB) de la Universidad Nacional Autónoma de México. Este autor agradece a Conacyt por los fondos recibidos durante sus estudios de Posgrado (160287) y a la Dirección General del Personal Académico (DGAPA) por el apoyo para realizar estudios de Posgrado.
También se agradece a Silvia Aguilar por revisar una primera versión de este manuscrito, a Julio César Montero-Rojas por el arreglo digital de las figuras y a 2 revisores anónimos, así como al editor asociado por los atinados comentarios que enriquecieron el manuscrito.
Material examinado de los géneros del complejo Milla. De los ejemplares del colector Gutiérrez se recolectaron muestras de 5 individuos por sitio, pero a los ejemplares de respaldo se les asignó un número de colecta por sitio, acrónimos de los herbarios revisados o donde se depositaron los ejemplares de acuerdo con Holmgren et al. (1990). Los ejemplares utilizados para el estudio anatómico se marcan con asterisco
Bessera elegans Schultes f. Estado de México: municipio Tejupilco, J. Gutiérrez y R. Ríos 1337, 1338* (FEZA, MEXU). Guerrero: municipio Coyuca de Catalán, J. Gutiérrez et al., 1242, 1243* (FEZA, MEXU); municipio Tlapehuala, J. Gutiérrez et al., 1236 (FEZA-MEXU). Jalisco: municipio Cocula, J. Gutiérrez et al., 1253* (FEZA-MEXU). Morelos: municipio Jonacatepec, J. Gutiérrez 1136 (FEZA, MEXU). Guerrero: municipio Arcelia, J. Gutiérrez 1237 (FEZA, MEXU). Nayarit: municipio El Nayar, J. Gutiérrez y E. Bravo 1386*, 1396* (FEZA, MEXU). Bessera tuitensis R. Delgadillo Jalisco: municipio Cabo Corrientes R. Ríos y J. Gutiérrez 997* (FEZA-MEXU); municipio La Huerta, J. Gutiérrez et al., 1252* (FEZA, MEXU). Bessera tenuiflora (Greene) Macbride Baja California Sur: municipio La Paz. J. L. León 7516* (HCIB, MEXU). Dandya balsensis López-Ferr. y Espejo Morelos: municipio Tlaquiltenango, J. Gutiérrez 791*, 797, 839, 1115* (FEZA). Dandya hannibalii L. W. Lenz Michoacán: municipio Huetamo, J. Gutiérrez et al., 1241* (FEZA, MEXU). Dandya purpusii H. E. Moore Coahuila: municipio Ramos Arizpe, J. Gutiérrez et al., 1225* (FEZA, MEXU). Dandya thadhowardii L. W. Lenz Guerrero: municipio Iguala, J. Gutiérrez 798*, 840, 841* (FEZA), J. Gutiérrez 840 (FEZA); municipio Coyuca de Catalán, J. Gutiérrez 804, 844* (FEZA). Michoacán: municipio San Lucas, J. Gutiérrez et al., 1225 (FEZA, MEXU); municipio Huetamo, J. Gutiérrez et al., 1240* (FEZA, MEXU). Milla biflora Cav. Chihuahua: municipio Basaseachi, J. Gutiérrez et al., 909* (FEZA). Chiapas: municipio Ocozocoautla, M. Ayala et al., 1000, 1012 (FEZA, MEXU); municipio Tuxtla Gutiérrez, M. Ayala et al., 978* (FEZA, MEXU); Distrito Federal: Delegación Coyoacán, J. Gutiérrez 1195* (FEZA, MEXU). Estado de México: municipio Atlacomulco, J. Gutiérrez et al., 1361* (FEZA, MEXU); municipio Bejucos, J. Gutiérrez y R. Ríos 1340* (FEZA, MEXU); municipio Huehuetoca, J. Gutiérrez y R. Ríos-Gómez 1131* (FEZA, MEXU); municipio Ixtapaluca, J. Gutiérrez et al., 1279 (FEZA, MEXU); municipio El Oro, J. Gutiérrez et al., 1362* (FEZA, MEXU); municipio Los Reyes La Paz, J. Gutiérrez 1149* (FEZA, MEXU); municipio Nepantla, J. Gutiérrez 1139* (FEZA, MEXU). Guanajuato: municipio San Luis de la Paz, J. Gutiérrez y R. Ríos-Gómez 1124*, 1127*, 1253-A (FEZA, MEXU); municipio San José Iturbide, J. Gutiérrez y R. Ríos-Gómez 1129*, 1169 (FEZA, MEXU). Guerrero: municipio Atenango del Río, J. Gutiérrez 1176* (FEZA, MEXU); municipio Xochipala, J. Gutiérrez y R. Ríos 1343, 1344* (FEZA, MEXU). Hidalgo: municipio Singuilican, J. Gutiérrez et al., 1165* (FEZA, MEXU). Jalisco: S/N C. Castillejos* (FEZA, MEXU). Michoacán: municipio Maravatio, J. Gutiérrez et al., 1365* (FEZA, MEXU); Morelos: municipio Jantetelco, J. Gutiérrez 1138* (FEZA, MEXU); municipio Jonacatepec, J. Gutiérrez 846, 1135* (FEZA, MEXU). Nayarit: municipio El Nayar, J. Gutiérrez y E. Bravo 1387, 1388*, 1389, 1391*, 1392, 1394*, 1398, 1399* (FEZA, MEXU). Oaxaca: Dto. Cuicatlán, J. Gutiérrez et al., 1336* (FEZA, MEXU); Dto. Juchitán, municipio Asunción Ixcaltepec J. Gutiérrez et al., 1357* (FEZA, MEXU); Dto. Miahuatlán, municipio Cuixtla, J. Gutiérrez et al., 1314* (FEZA, MEXU); municipio Amatlán, J. Gutiérrez et al., 1315, 1316* (FEZA, MEXU); Dto. Tlaxiaco, municipio Tlaxiaco, J. Gutiérrez et al., 1154 *, 1156 (FEZA, MEXU); Dto. Juxtlahuaca, municipio Tecomaxtlahuaca, J. Gutiérrez et al., 1160* (FEZA, MEXU); Dto. Putla, municipio Putla, J. Gutiérrez et al., 1173* (FEZA, MEXU); Dto. Huajuapan municipio Santo Domingo Tonalá J. Gutiérrez y R. Ríos 1271* (FEZA, MEXU); municipio San Marcos de Arteaga J. Gutiérrez et al.1162* (FEZA y MEXU). Dto. Yautepec, municipio Nejapa de Madero, J. Gutiérrez et al., 1356* (FEZA, MEXU); municipio Santiago Chazumba, R. Ríos y J. Gutiérrez 896 (FEZA, MEXU); municipio San Juan Bautista Suchitepec, R. Ríos y J. Gutiérrez 898 (FEZA, MEXU); municipio Zapotitlán Palmas, R. Ríos y J. Gutiérrez 902 (FEZA, MEXU). Puebla: municipio Azumbilla, J. Gutiérrez y R. Ríos 1263* (FEZA, MEXU); municipio Tecamachalco, J. Gutiérrez y R. Ríos 1255* (FEZA, MEXU); municipio Tepeaca, J. Gutiérrez y R. Ríos 1254* (FEZA, MEXU); municipio Tehuacán, J. Gutiérrez y R. Ríos 1267*, 1268* (FEZA, MEXU); J. Gutiérrez et al., 1354* (FEZA, MEXU); municipio Tlacotepec, J. Gutiérrez y R. Ríos 1256*, 1257* (FEZA, MEXU); municipio Toltepec de Guerrero, J. Gutiérrez y R. Ríos 1274*, 1275*, 1276* (FEZA, MEXU). Querétaro: municipio Cadereyta, J. Gutiérrez y R. Ríos-Gómez 1130* (FEZA, MEXU); municipio Jalpan de Serra, J. Gutiérrez 1196* (FEZA, MEXU). San Luis Potosí: municipio Santa María del Río, J. Gutiérrez 1210* (FEZA, MEXU); municipio Zaragoza, R. Ríos 898 (FEZA, MEXU). Veracruz: municipio Maltrata, J. Gutiérrez et al., 1304* (FEZA, MEXU). Zacatecas: municipio Valparaíso, J. Gutiérrez y E. Bravo 1400*, 1401*, 1402*, 1403*, 1404*, 1405, 1406* (FEZA, MEXU); municipio Zacatecas, J. Gutiérrez y E. Bravo 1407 (FEZA, MEXU). Milla bryani I. M. Johnst. Coahuila: municipio Cuatro Ciénegas, J. Gutiérrez et al., 1226* (FEZA, MEXU). Milla filifolia T. M. Howard Morelos: municipio Tenango, J. Gutiérrez 1133* (FEZA, MEXU). Milla magnifica H. E. Moore. Guerrero: municipio Iguala, J. Gutiérrez y R. Muñoz 823 (FEZA), J. Gutiérrez 838 (FEZA); municipio Taxco, J. Gutiérrez et al., 1235 (FEZA, MEXU); municipio Teloloapan, J. Gutiérrez e I. Escalante 812* (FEZA). Milla mexicana T. M. Howard Puebla: municipio Izúcar de Matamoros, J. Gutiérrez 1134* (FEZA, MEXU); municipio Huehuetlán el Chico, J. Gutiérrez 1137 (FEZA, MEXU). Milla mortoniana H. E. Moore Michoacán: municipio Aquila, Sánchez-Mejorada et al., 4301 (MEXU). Milla oaxacana Ravenna. Oaxaca: Dto. Cuicatlán, municipio Santiago Nacaltepec, J. Gutiérrez et al., 1327 (FEZA, MEXU); municipio Oaxaca de Juárez, J. Gutiérrez et al., 1185, 1186 (FEZA, MEXU); Dto. Etla, municipio San Pablo Huitzo, J. Gutiérrez et al., 1192, 1193* (FEZA, MEXU). Milla potosina T. M. Howard San Luis Potosí: municipio Guadalcazar, J. Gutiérrez e I. Escalante 1414* (FEZA, MEXU). Milla rosea H. E. Moore Nuevo León: municipio Montemorelos, J. Gutiérrez y E. Estrada 1218* (FEZA, MEXU). Milla valliflora J. Gut. y E. Solano Dto. Nochixtlán, municipio Suchixtlahuaca, J. Gutiérrez et al., 1150, 1151* (FEZA, MEXU). Jaimehintonia gypsophila Turner Nuevo León: municipio Aramberri, J. Gutiérrez y E. Estrada 1204, 1208* (FEZA, MEXU). Petronymphe decora H. E. Moore Guerrero: municipio Chilpancingo de los Bravo, J. Gutiérrez y R. Ríos 1350* (FEZA, MEXU).
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.