








Con el fin de actualizar y resumir la información sobre la diversidad de los medusozoarios presentes en aguas mexicanas, se realizó una revisión por familia de los registros disponibles para establecer el estado actual de nuestro conocimiento de la biodiversidad nacional de este grupo. Se presenta una revisión sintética de diversos aspectos de su biología y ecología. Además, se incluyen nuevos registros de: Crossota millsae, Halitrephes maasi, Atorella octogonos, Tiburonia granrojo, Stygiomedusa gigantea, Periphyllopsis braweri, Resomia persica, Bargmannia lata, Desmophyes haematogaster y Lensia quadriculata, especies encontradas en el golfo de California. Los resultados de este análisis muestran un total de 289 especies de Medusozoa reconocidas en México, de las cuales 3 son de Cubozoa, 20 de Scyphozoa y 266 de Hydrozoa. En total, el número de especies registradas en aguas mexicanas representa el 7.3% de los medusozoarios conocidos actualmente en el mundo, lo que indica que la diversidad de este grupo de organismos merece mayores esfuerzos, particularmente en ambientes poco explorados.
In order to update and summarize the information about the diversity of the medusozoans occurring in Mexican waters, records were reviewed at the family level to assess the current status of the knowledge of the diversity of this group in Mexico. A synthesized revision of different biological and ecological aspects of the group is also presented. In addition, new records are included of: Crossota millsae, Halitrephes maasi, Atorella octogonos, Tiburonia granrojo, Stygiomedusa gigantea, Periphyllopsis braweri, Resomia persica, Bargmannia lata, Desmophyes haematogaster and Lensia quadriculata, found in the Gulf of California. The results of this analysis yielded a total of 289 species of Medusozoa known in Mexico, of which 3 species are of Cubozoa, 20 of Scyphozoa, and 266 of Hydrozoa. Overall, the number of species recorded in Mexican waters represents about 7.3% of the medusozoans known worldwide, thus suggesting that the diversity of the group deserves further efforts, particularly in scarcely surveyed habitats.
Los medusozoos o medusozoarios (medusas) incluyen tres grandes grupos, los Cubozoa, los Scyphozoa y los Hydrozoa que junto con los Anthozoa (corales y anémonas) forman el phylum Cnidaria. Los medusozoos son uno de los componentes más importantes de las comunidades bióticas marinas y sus integrantes forman parte tanto del zooplancton, representados básicamente por medusas y sifonóforos, como del bentos, representados por los pólipos y diferentes estadios de resistencia. Debido a las características de su ciclo de vida, tienen la capacidad de aprovechar condiciones locales para producir floraciones con dramáticos incrementos de su abundancia. Estos episodios causan efectos disruptivos en las comunidades biológicas de la columna de agua (Mills, 2001; Purcell et al., 2007). Aún no se conocen con precisión las causas de tales florecimientos, y aunque no hay consenso, se han sugerido como causales potenciales a la variabilidad en los ecosistemas, el calentamiento global, perturbaciones antropogénicas y la eliminación de depredadores tope en las tramas tróficas (Boero et al., 2008).
Desde el punto de vista taxonómico, las clases y otros niveles establecidos en las clasificaciones actuales de los cnidarios no tienen relación con su filogenia (Daly et al., 2007), y debido a las diferentes interpretaciones de su genética, morfología y ciclo de vida, no hay un acuerdo entre los especialistas en cuanto a la adopción de una clasificación única (Bouillon et al., 2006; Collins et al., 2006; Daly et al., 2007). Es por ello que en este trabajo se tratan simplemente como grupos sin especificar categorías taxonómicas suprafamiliares.
Los estudios generales o de revisión más importantes son los de Bouillon (1999), Bouillon y Boero (2000), Bouillon et al. (2006) y Daly et al. (2007) para las medusas y los de Totton (1965) y Pugh (1999) para los sifonóforos, aunque después de éstos últimos se han publicado varias revisiones de distintos taxa. En el presente trabajo se actualiza la información sobre la diversidad de los medusozoarios presentes en aguas mexicanas.
Descripción morfológica de MedusozoaTodos los cnidarios son metazoarios acelomados, diploblásticos, con nivel de construcción tisular, sin órganos como tales y comparten la presencia de organelos subcelulares altamente complejos llamados cnidoblastos (=cnidocistos) (cápsulas urticantes o adhesivas) del tipo nematocisto (urticantes) que pueden ser de distintas clases. La mayoría de ellos funcionan para inyectar toxina y envenenar a la presa o al agresor mediante la proyección de largos filamentos tubulares perforantes huecos; los filamentos de tipo adhesivo tienen como función enredarse o adherirse a sustratos. De acuerdo al grupo en el que se presentan se les denomina nematocistos, spirocistos (exclusivos de los Anthozoa) y pticocistos (restringidos a los Anthozoa del orden Cerianthara); en conjunto hay más de 30 variedades (Mariscal, 1974; Kass-Simon y Scappaticci, 2002). Debido a sus mecanismos de descarga, penetración y de inyección de toxinas, los cnidocistos se encuentran entre las estructuras subcelulares más complejas de cualquier phylum conocido (Williamson et al., 1996). La gran sobrevivencia y éxito de los cnidarios puede ser el resultado, en gran medida, de la evolución de los nematocistos y de sus potentes toxinas.
Los cnidarios son organismos simples anatómica y fisiológicamente. Su sistema digestivo o celenterón con forma de saco simple o ramificado, tiene sólo un orificio que sirve como boca y ano. Generalmente tienen tentáculos cubiertos con cnidocistos. Los medusozoos se distinguen de los antozoos en que estos últimos sólo presentan pólipos (=hydroides) y los primeros desarrollan además una forma medusa libre nadadora como parte de su ciclo de vida. Las medusas presentan una gran diversidad de formas y tienen alternancia de generaciones (ciclo metagenético) en la que típicamente, aunque no siempre, un pólipo béntico asexual se alterna con una medusa planctónica sexual. La medusa puede estar reducida a gonóforos que no abandonan la colonia hydroide y en otros casos la medusa puede producir directamente otra medusa. El máximo grado de polimorfismo es alcanzado en hidrozoos del grupo Siphonophora.
Los hydroides pueden ser solitarios, pero generalmente forman colonias por gemación. El brote asexual de nuevas medusas o gonóforos generalmente involucra la formación de un nódulo medusario o ectocodon. Los individuos de las colonias están interconectados al celenterón y pueden producir pólipos especializados en diferentes funciones; así, los dactilozooides son individuos defensivos, los gonozoides reproductivos, los gastrozoides nutricionales, etc. Las medusas son generalmente carnívoras, aunque algunas se alimentan de bacterias, protozoarios, algas, fitoplancton o materia orgánica disuelta.
La forma pólipo es bentónica, sésil y generalmente se encuentra en agregaciones de organismos similares (Fig. 1). Los hydrantes o pólipos alimentadores pueden tener varias formas, tales como urna, clava, cónica, cilíndrica, etc. El ápice de los hidrantes forma una probosis o hypostoma cónico o en forma de domo, con una boca terminal. Por debajo del hypostoma puede haber tentáculos; ambas estructuras son muy importantes en la alimentación. Los tentáculos son las estructuras más característica de los hydrantes y varían en tipo y estructura de acuerdo a la manera en la que los cnidocistos están distribuidos en su superficie.

La columna gástrica es un alargamiento simple y no dividido de la parte digestiva del hydrante. Puede poseer tentáculos y brotes medusarios y gonóforos. Los tentáculos pueden ser sólidos o tener una extensión del canal gastrovascular, generalmente son muy contráctiles y tienen las funciones de defensa, alimentación y en muchos casos, de anclaje. Los tentáculos se clasifican de acuerdo a la distribución de los cnidocistos en su superficie.
La forma medusa es planctónica, solitaria y libre nadadora y típicamente tiene simetría tetrámera, o polímera (Fig. 1). El cuerpo tiene forma de hongo, campana o disco. La mesoglea o material gelatinoso de la medusa le da forma y favorece su flotación. El exterior de la medusa, convexa y aboral es la exumbrela y la parte interna, cóncava y oral es la subumbrela, la cual puede ser angostada por un diafragma muscular horizontal o velo, primordial para el desplazamiento de la medusa. El borde de la umbrela tiene tentáculos (marginales), ocelos, estatocistos y cordili. Los tentáculos pueden disponerse también alrededor de la boca (tentáculos orales). El manubrio es una proyección de la subumbrela que tiene en posición distal la cavidad gástrica; proximalmente se extiende en los canales radiales que sirven para distribuir el alimento, los desechos, y los cnidoblastos hasta los gametos. El manubrio puede tener tamaño y formas muy variables. La boca puede ser simple, o con labios, lóbulos o tentáculos.
Las células sexuales pueden madurar en el manubrio, en los canales radiales, o en ambos. La fertilización es generalmente externa. El embrión se desarrolla en una larva plánula que se asienta para a su vez transformarse en un nuevo pólipo o en ocasiones en una nueva medusa.
CubozoaEn particular, los Cubozoa o medusas en forma de caja constituyen un grupo (Cubomedusae) distintivo que representan, por su picadura, un peligro para el ser humano. Entre las cubomedusas es notoria la avispa de mar australiana (Chironex fleckeri) cuya toxina puede ser fatal al igual que las llamadas irukandji, como Malo kingi y Carukia barnesi. Son medusas de forma cúbica, con cnidoblastos especiales, muy potentes. Hasta hace poco se le consideraba como un orden de los Scyphozoa. Se caracterizan por su división tetrámera, que separa el celenterón en cuatro bolsas. Presentan cuatro ropalia sensoriales perradiales con ojos y ocelos, cuerpos vítreos, lentes y retinas, además de estatocistos. El borde umbrelar no es festoneado y el margen de la subumbrela se pliega al interior para formar un velario que angosta la apertura subumbrelar. Los tentáculos o grupos de tentáculos se originan en cada una de las cuatro esquinas interradiales y tienen bases musculares engrosadas llamadas pedalia. Uno de los aspectos más particulares de las cubomedusas es que poseen un ojo complejo asociado a cada ropalio que les permite seguir objetos en movimiento y responder rápidamente a cambios de intensidad de luz.
ScyphozoaLas escifomedusas están representadas por medusas de talla grande que incluyen a la mayoría de los más grandes invertebrados marinos solitarios. Los episodios de enjambre de estas medusas suelen afectar la dinámica del ambiente marino. Aún así, y paradójicamente, para la mayoría de las regiones del mundo no existe un recuento taxonómico confiable de ellas y su posición y patrones biológicos en las comunidades marinas aún no se entienden por completo.
Las escifomedusas poseen una mesoglea o capa intermedia relativamente gruesa que les da un mayor soporte corporal. Presentan simetría radial y se considera que las escifomedusas son los organismos no bilaterales más perfectamente simétricos y de mayor tamaño conocidos en el planeta, lo que les proporciona un notable atractivo estético (Fig. 2).
.")
La anatomía de las escifomedusas es similar a la de Hydrozoa, que en general es simple. Son acraspédotas, es decir, carecen de velo. Presentan cnidocistos en la cavidad gastrovascular. Las gónadas son endodérmicas; una vez maduras, pueden almacenarse en la endodermis. Su cavidad gastrovascular está dividida de manera incompleta por 4 tabiques o septos parciales en posición inter-radial. Los tabiques separan 4 bolsas gástricas.
Las escifomedusas contienen distintos grupos: Stauromedusae, la mayoría son formas pequeñas, con una fase polipoide; Coronatae, grupo representado por medusas de aguas meso y batipelágicas que poseen un surco profundo (surco coronal) que divide a la exumbrela en 2 regiones, interna y externa; Semaestomeae, medusas típicamente grandes, conspicuas, con forma de disco invertido y que son más abundantes en aguas someras costeras; y Rhizostomae medusas típicamente hemisféricas, de talla mediana a grande que en su mayoría habitan aguas cálidas costeras. Contiene formas filtradoras y la mayoría de las especies se distinguen por tener numerosas bocas localizadas en los brazos orales en vez de una sola boca central.
HydrozoaLos Hydrozoa conforman un grupo heterogéneo que incluye a los cnidarios que producen un estadio de vida libre pelágico que se reproduce sexualmente y que se conoce como medusa (Figs. 3, 4). Comparten también la presencia de velo y gónadas ectodérmicas (Boero y Bouillon, 2000). El grupo se ha subdividido en 3 categorías con base en la forma en que las medusas se desarrollan a partir de los pólipos. En los Automedusa la medusa se produce por la transformación directa de la plánula (nunca béntica) o a través de un “estado“ actínula. En los Hydroidomedusa las medusas se producen a través de un nódulo medusario y en los Polypodiozoa este proceso se genera mediante un complejo patrón de desarrollo parasitario (Bouillon et al., 2006). En las medusas se reconocen distintos tipos de tentáculos que se clasifican según la distribución de los cnidocistos.
.")
.")
En las cubomedusas el tamaño generalmente está entre 2 y 4cm pero pueden llegar a alcanzar entre 15 y 25cm; tienden a ser transparentes, lo que hace difícil verlas en el agua incluso en grandes números. Las escifomedusas pueden medir desde unos cuantos milímetros hasta cerca de 2m. Los géneros Desmonema y Chrysaora pueden alcanzar más de 1m de diámetro umbrelar y en Cyanea se han reportado individuos de hasta 2m de diámetro y varios metros de largo. Muchas especies están profunda y brillantemente coloreadas, con patrones de color distribuidos radialmente, lo que añade belleza al grupo; sin embargo, esta coloración se pierde o debilita después de la fijación.
Los Hydrozoa son generalmente formas pequeñas que miden algunos centímetros (0.5-6cm). Una excepción es el diverso grupo de los sifonóforos (Fig. 5), cuyas colonias pueden medir decenas de metros, por sus largos tentáculos. Los organismos pueden estar coloreados, principalemente los que viven en la zona oscura del océano.
.")
La bioluminiscencia es otra característica de escifo e hidromedusas, pero no de cubomedusas, y les sirve principalmente como defensa o advertencia, pero también, algunos sifonóforos la usan como carnada para atraer a sus presas (Haddock et al., 2010).
HábitatLas cubo y escifomedusas han colonizado una gran diversidad de hábitats, poseen amplios rangos de distribución abarcando la plataforma continental y el área oceánica. El endemismo es raro en ambos grupos y muchas especies son consideradas cosmopolitas. La distribución vertical de ambos grupos está definida por una conjunción de factores como la luz, presión, presencia de presas, etapa en el ciclo de vida, así como gradientes de temperatura, salinidad y oxígeno. Las escifomedusas son exclusivamente marinas y algunas de ellas son capaces de realizar extensos desplazamientos verticales (p.ej. Periphylla periphylla y Atolla vanhoeffeni).
Dentro de los Hydrozoa, las automedusas son principalmente marinas, solitarias y pelágicas. Aún cuando el grupo es extremadamente exitoso en el ámbito marino, existen algunos representantes dulceacuícolas, menos de 40 especies, en todos los ambientes acuáticos continentales desde mesotróficos hasta eutróficos (Jankowski et al., 2008). Las especies dulceacuícolas comprenden 4 grupos filogenéticamente dispersos: 1) la común hidra (Hydra), 2) los Cordylophorinae anthoatecados con hidroides coloniales, 3) las medusas de agua dulce Craspedacusta y Limnocnida y 4) Polypodium, un inusual parásito de huevos de peces asignado a una clase propia: Polypodioza (Boero y Bouillon, 2000). Las Hydroidomedusa generalmente alternan un estadio pólipo béntico y sésil con un estadio medusa pelágico, de vida libre.
Ciclos de vidaCubozoa. Algunas especies de cubomedusas de tamaño grande viven varios años, pero la mayoría vive algunas semanas o meses. Presentan una gran capacidad de dispersión, ya que pueden desplazarse en amplias áreas geográficas, por cientos o miles de kilómetros desde su área de origen. Todas las cubomedusas cuyo ciclo de vida es conocido poseen una etapa de pólipo. Los pólipos de las cubomedusas son muy pequeños y se pueden reproducir asexualmente mediante bipartición para formar nuevos pólipos. Sin embargo, no estrobilan, por el contrario, se desarrollan directamente por metamorfosis en una pequeña medusa. La acumulación en enjambres de cubomedusas está en gran medida en función de los procesos reproductivos de cópula, único dentro del phylum.
ScyphozoaLa reproducción de las escifomedusas es estacional, excepto en latitudes tropicales. Aún cuando la mayoría de las especies son dioicas, algunas pocas son hermafroditas típicamente siendo protándricas o protogínicas. Algunas son ligeramente paedomórficas, maduran sexualmente antes que todos sus caracteres sexuales de adulto se han desarrollado (p. ej. Chrysaora). Algunas escifomedusas de aguas profundas pueden vivir varios años pero la mayoría vive pocas semanas o meses. Típicamente los ciclos de vida incluyen una etapa polipoide béntica, llamada escifistoma. Las pequeñas larvas medusoides o éfiras, estrobilan (se desprenden) a partir de la parte superior del escifistoma. Casi todas las escifomedusas alternan una etapa béntica asexual sésil (escifistoma) y una planctónica sexual (medusa). Cada éfira se desarrolla en una nueva medusa que puede ser hembra o macho. Un sólo escifistoma puede producir algunas a muchas medusas. Algunas escifistomas pueden vivir varios años por lo que parece ser un patrón común y, durante sus vidas, pueden liberar cientos de éfiras. Algunas escifomedusas de tallas grandes pudieron ser liberadas cientos o tal vez miles de kilómetros lejos del área en la que comenzaron su etapa reproductiva. Esto significa que es probable que se haya dado un intercambio genético panoceánico; la mayoría de las especies están distribuidas en extensas áreas geográficas. La mayoría de las escifomedusas con ciclos de vida conocidos poseen una etapa de pólipo. Se conocen los ciclos de vida de algunas especies costeras pero para las demás especies, que aún no han sido estudiadas, la etapa bentónica permanece desconocida. En algunas escifomedusas Coronatae la etapa medusa es vestigial o ausente, una tendencia evolutivamente paralela y común que se ha documentado en los Hydrozoa Leptolida. En contraste, en al menos una especie de Pelagia, la etapa del escifistoma se ha suprimido en su ciclo de vida, lo que le permite distribuirse en mar abierto.
HydrozoaLos ciclos de vida de los Hydrozoa son los más complejos y variados del reino animal y son poco conocidos, pero se pueden distinguir 4 tipos principales (Bouillon et al., 2006): a) automedusa, en el que los embriones o plánula nunca son bénticos y se desarrollan directamente o por medio de larvas actinuloides pelágicas en una medusa adulta sexual; b) el de la mayoría de los Hydroidomedusa en el que la plánula se desarrolla en un estadio larvario, los pólipos, esencialmente bénticos, y que pueden ser solitarios o agruparse, y que posteriormente engendrarán por reproducción asexual, vía un nódulo medusario, al estadio de medusa adulto, sexual, solitario y pelágico. Este ciclo puede tener numerosas variantes y la más significativa es la supresión de la medusa. Este ciclo es compartido por cerca de la mitad de las especies de hidroidomedusas. En éste, el estadio medusa es reducido en grados variables a esporosacos (gonóforos) fijos a pólipos y de los que se diferencian los gametos. Los pólipos larvales producen por pedogénesis el estadio adulto sexuado; c) el tipo hidroidomedusa Siphonophora, donde las plánulas no se fijan nunca y se desarrollan en colonias pelágicas, extremadamente polimórficas compuestas de diferentes tipos de polipoides y donde las medusas están reducidas a órganos flotadores o propulsores y d) el de los Polipodiozoa, estadios endocelulares parásitos (polipoides?) que producen estadios tentaculados de vida libre (medusoides?).
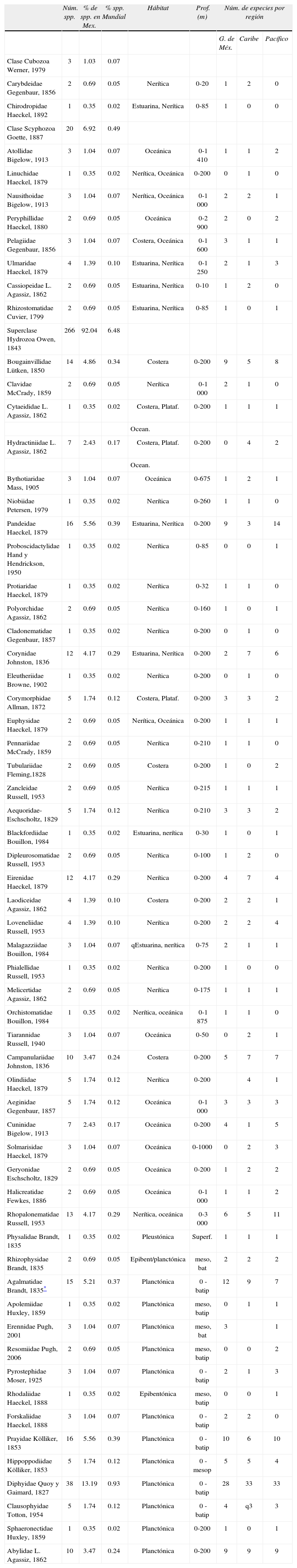
DiversidadEl número de especies de Medusozoa distribuidas en México es de 289 (Cuadro 1) y pertenecen a las clases Cubozoa (2 familias, 3 especies), Scyphozoa (7 familias, 20 especies) y a la superclase Hydrozoa (52 familias y 266 especies) de los cuales 106 especies pertenecen a la subclase Siphonophorae. La mayoría de las especies registradas son formas nerítico-costeras, excepto los sifonóforos (Fig. 5) que son generalmente holoplanctónicos y oceánicos; pocas especies, relativamente, son completamente oceánicas o estuarinas. Ésto puede estar relacionado a esfuerzos de muestreo más intensos en las áreas costeras y de plataforma, principalmente en la columna de agua (Segura-Puertas et al., 2003, 2010; Mendoza-Becerril et al., 2009).
Número de especies de Medusozoa registradas en México, sumadas por familia, con el porcentaje de especies que representan para México y para el número de especies conocidas en el mundo y datos de su hábitat y distribución
| Núm. spp. | % de spp. en Mex. | % spp. Mundial | Hábitat | Prof. (m) | Núm. de especies por región | |||
| G. de Méx. | Caribe | Pacífico | ||||||
| Clase Cubozoa Werner, 1979 | 3 | 1.03 | 0.07 | |||||
| Carybdeidae Gegenbaur, 1856 | 2 | 0.69 | 0.05 | Nerítica | 0-20 | 1 | 2 | 0 |
| Chirodropidae Haeckel, 1892 | 1 | 0.35 | 0.02 | Estuarina, Nerítica | 0-85 | 1 | 0 | 0 |
| Clase Scyphozoa Goette, 1887 | 20 | 6.92 | 0.49 | |||||
| Atollidae Bigelow, 1913 | 3 | 1.04 | 0.07 | Oceánica | 0-1 410 | 1 | 1 | 2 |
| Linuchidae Haeckel, 1879 | 1 | 0.35 | 0.02 | Nerítica, Oceánica | 0-200 | 0 | 1 | 0 |
| Nausithoidae Bigelow, 1913 | 3 | 1.04 | 0.07 | Nerítica, Oceánica | 0-1 000 | 2 | 2 | 1 |
| Peryphillidae Haeckel, 1880 | 2 | 0.69 | 0.05 | Oceánica | 0-2 900 | 2 | 0 | 2 |
| Pelagiidae Gegenbaur, 1856 | 3 | 1.04 | 0.07 | Costera, Oceánica | 0-1 600 | 3 | 1 | 1 |
| Ulmaridae Haeckel, 1879 | 4 | 1.39 | 0.10 | Estuarina, Nerítica | 0-1 250 | 2 | 1 | 3 |
| Cassiopeidae L. Agassiz, 1862 | 2 | 0.69 | 0.05 | Estuarina, Nerítica | 0-10 | 1 | 2 | 0 |
| Rhizostomatidae Cuvier, 1799 | 2 | 0.69 | 0.05 | Estuarina, Nerítica | 0-85 | 1 | 0 | 1 |
| Superclase Hydrozoa Owen, 1843 | 266 | 92.04 | 6.48 | |||||
| Bougainvillidae Lütken, 1850 | 14 | 4.86 | 0.34 | Costera | 0-200 | 9 | 5 | 8 |
| Clavidae McCrady, 1859 | 2 | 0.69 | 0.05 | Nerítica | 0-1 000 | 2 | 1 | 0 |
| Cytaeididae L. Agassiz, 1862 | 1 | 0.35 | 0.02 | Costera, Plataf. | 0-200 | 1 | 1 | 1 |
| Ocean. | ||||||||
| Hydractiniidae L. Agassiz, 1862 | 7 | 2.43 | 0.17 | Costera, Plataf. | 0-200 | 0 | 4 | 2 |
| Ocean. | ||||||||
| Bythotiaridae Mass, 1905 | 3 | 1.04 | 0.07 | Oceánica | 0-675 | 1 | 2 | 1 |
| Niobiidae Petersen, 1979 | 1 | 0.35 | 0.02 | Nerítica | 0-260 | 1 | 1 | 0 |
| Pandeidae Haeckel, 1879 | 16 | 5.56 | 0.39 | Estuarina, Nerítica | 0-200 | 9 | 3 | 14 |
| Proboscidactylidae Hand y Hendrickson, 1950 | 1 | 0.35 | 0.02 | Nerítica | 0-85 | 0 | 0 | 1 |
| Protiaridae Haeckel, 1879 | 1 | 0.35 | 0.02 | Nerítica | 0-32 | 1 | 1 | 0 |
| Polyorchidae Agassiz, 1862 | 2 | 0.69 | 0.05 | Nerítica | 0-160 | 1 | 0 | 1 |
| Cladonematidae Gegenbaur, 1857 | 1 | 0.35 | 0.02 | Nerítica | 0-200 | 0 | 1 | 0 |
| Corynidae Johnston, 1836 | 12 | 4.17 | 0.29 | Estuarina, Nerítica | 0-200 | 2 | 7 | 6 |
| Eleutheriidae Browne, 1902 | 1 | 0.35 | 0.02 | Nerítica | 0-200 | 0 | 1 | 0 |
| Corymorphidae Allman, 1872 | 5 | 1.74 | 0.12 | Costera, Plataf. | 0-200 | 3 | 3 | 2 |
| Euphysidae Haeckel, 1879 | 2 | 0.69 | 0.05 | Nerítica, Oceánica | 0-200 | 1 | 1 | 1 |
| Pennariidae McCrady, 1859 | 2 | 0.69 | 0.05 | Nerítica | 0-210 | 1 | 1 | 0 |
| Tubulariidae Fleming,1828 | 2 | 0.69 | 0.05 | Costera | 0-200 | 1 | 0 | 2 |
| Zancleidae Russell, 1953 | 2 | 0.69 | 0.05 | Nerítica | 0-215 | 1 | 1 | 1 |
| Aequoridae-Eschscholtz, 1829 | 5 | 1.74 | 0.12 | Nerítica | 0-210 | 3 | 3 | 2 |
| Blackfordiidae Bouillon, 1984 | 1 | 0.35 | 0.02 | Estuarina, nerítica | 0-30 | 1 | 0 | 1 |
| Dipleurosomatidae Russell, 1953 | 2 | 0.69 | 0.05 | Nerítica | 0-100 | 1 | 2 | 0 |
| Eirenidae Haeckel, 1879 | 12 | 4.17 | 0.29 | Nerítica | 0-200 | 4 | 7 | 4 |
| Laodiceidae Agassiz, 1862 | 4 | 1.39 | 0.10 | Costera | 0-200 | 2 | 2 | 1 |
| Loveneliidae Russell, 1953 | 4 | 1.39 | 0.10 | Nerítica | 0-200 | 2 | 2 | 4 |
| Malagazziidae Bouillon, 1984 | 3 | 1.04 | 0.07 | qEstuarina, nerítica | 0-75 | 2 | 1 | 1 |
| Phialellidae Russell, 1953 | 1 | 0.35 | 0.02 | Nerítica | 0-200 | 1 | 0 | 0 |
| Melicertidae Agassiz, 1862 | 2 | 0.69 | 0.05 | Nerítica | 0-175 | 1 | 1 | 1 |
| Orchistomatidae Bouillon, 1984 | 1 | 0.35 | 0.02 | Nerítica, oceánica | 0-1 875 | 1 | 1 | 0 |
| Tiarannidae Russell, 1940 | 3 | 1.04 | 0.07 | Oceánica | 0-50 | 0 | 2 | 1 |
| Campanulariidae Johnston, 1836 | 10 | 3.47 | 0.24 | Costera | 0-200 | 5 | 7 | 7 |
| Olindiidae Haeckel, 1879 | 5 | 1.74 | 0.12 | Nerítica | 0-200 | 4 | 1 | |
| Aeginidae Gegenbaur, 1857 | 5 | 1.74 | 0.12 | Oceánica | 0-1 000 | 3 | 3 | 3 |
| Cuninidae Bigelow, 1913 | 7 | 2.43 | 0.17 | Oceánica | 0-200 | 4 | 1 | 5 |
| Solmarisidae Haeckel, 1879 | 3 | 1.04 | 0.07 | Oceánica | 0-1000 | 0 | 2 | 3 |
| Geryonidae Eschscholtz, 1829 | 2 | 0.69 | 0.05 | Oceánica | 0-200 | 1 | 2 | 2 |
| Halicreatidae Fewkes, 1886 | 2 | 0.69 | 0.05 | Oceánica | 0-1 000 | 1 | 1 | 2 |
| Rhopalonematidae Russell, 1953 | 13 | 4.17 | 0.29 | Nerítica, oceánica | 0-3 000 | 6 | 5 | 11 |
| Physalidae Brandt, 1835 | 1 | 0.35 | 0.02 | Pleustónica | Superf. | 1 | 1 | 1 |
| Rhizophysidae Brandt, 1835 | 2 | 0.69 | 0.05 | Epibent/planctónica | meso, bat | 2 | 2 | 2 |
| Agalmatidae Brandt, 1835* | 15 | 5.21 | 0.37 | Planctónica | 0 - batip | 12 | 9 | 7 |
| Apolemiidae Huxley, 1859 | 1 | 0.35 | 0.02 | Planctónica | meso, batip | 0 | 1 | 1 |
| Erennidae Pugh, 2001 | 3 | 1.04 | 0.07 | Planctónica | meso, bat | 3 | 1 | |
| Resomiidae Pugh, 2006 | 2 | 0.69 | 0.05 | Planctónica | meso, batip | 0 | 0 | 2 |
| Pyrostephidae Moser, 1925 | 3 | 1.04 | 0.07 | Planctónica | 0 - batip | 2 | 1 | 3 |
| Rhodaliidae Haeckel, 1888 | 1 | 0.35 | 0.02 | Epibentónica | meso, batip | 0 | 0 | 1 |
| Forskaliidae Haeckel, 1888 | 3 | 1.04 | 0.07 | Planctónica | 0 - batip | 2 | 2 | 0 |
| Prayidae Kölliker, 1853 | 16 | 5.56 | 0.39 | Planctónica | 0 - batip | 10 | 6 | 10 |
| Hippoppodiidae Kölliker, 1853 | 5 | 1.74 | 0.12 | Planctónica | 0 - mesop | 5 | 5 | 4 |
| Diphyidae Quoy y Gaimard, 1827 | 38 | 13.19 | 0.93 | Planctónica | 0 - batip | 28 | 33 | 33 |
| Clausophyidae Totton, 1954 | 5 | 1.74 | 0.12 | Planctónica | 0 - batip | 4 | q3 | 3 |
| Sphaeronectidae Huxley, 1859 | 1 | 0.35 | 0.02 | Planctónica | 0-200 | 1 | 0 | 1 |
| Abylidae L. Agassiz, 1862 | 10 | 3.47 | 0.24 | Planctónica | 0-200 | 9 | 9 | 9 |
En México se han registrado 289 especies de medusozoarios desde 1897 (Fig. 6); en el Pacífico se han registrado 176 (61%) especies, 167 (58%) en las aguas del golfo de México y 161 (56%) en el Caribe mexicano. No hay evidencias de endemismo en estos grupos en México.

Mundialmente se conocen alrededor de 3 702 especies dentro de la superclase Hydrozoa (Bouillon et al., 2006). El grupo que posee menos especies es el de las automedusas (134), 3 567 pertenecen a las hidroidomedusas y sólo se conoce una especie de la clase Polipodiozoa (Bouillon et al., 2006) de la cual no se tienen registros para este grupo en México. Se conocen también 200 especies de escifomedusas (Mianzan y Cornelius, 1999) y 36 especies de cubomedusas (Daly et al., 2007), lo que hace un total de 3 938 especies registradas en el mundo del grupo Medusozoa. De ese total mundial, a México le corresponde el 7.3%, 289 especies.
Conforme se incrementen los trabajos que incluyan las etapas hidroides de pólipos bénticos y el análisis de las medusas liberadas en laboratorio a partir de éstos, se irán encontrando mayor número de especies aún no registradas para aguas mexicanas o se corregirán registros inválidos. Las regiones abisales o batiales y meso y batipelágicas, son ambientes diversos para algunos taxa del zooplancton, como los cnidarios y es de esperarse que se encuentren nuevas especies y nuevos registros a partir de muestras recolectadas en aguas profundas (Gasca, 2009). Se sabe ya que hay especies nuevas e incluso más de una docena de especies de sifonóforos ya recolectadas y en espera de ser descritas. Aún siguen siendo escasos los estudios sobre medusas en aguas tropicales del Pacífico mexicano; es más conocida la zona pelágica de esta última área y la zona de plataforma aún permanece prácticamente desconocida; es de esperarse que surjan más especies conforme se aumenten los estudios en esta vasta área.
Dentro de las especies más comunes y abundantes en el Pacífico y Atlántico mexicanos se encuentran: Bougainvillia muscus, Lizzia alvarinoae, Cytaeis tetrastyla, Stomotoca pterophylla, Euphysora gracilis, Eucheilota duodecimalis, Clytia discoida, C. folleata, Liriope tetraphylla, Solmundella bitentaculata, Cunina octonaria, Aglaura hemistoma, Rhopalonema velatum, Nausithöe punctata, Pelagia noctiluca y Aurelia aurita (Segura-Puertas et al., 2003, 2010; Mendoza-Becerril et al., 2009). Entre los sifonóforos más comúnmente recolectados en aguas epipelágicas están: Abylopsis eschscholtzi, Chelophyes appendiculata, Dyphies bojani, D. dispar, Eudoxoides spiralis y Bassia bassensis. En el Pacífico mexicano son abundantes Muggiaea atlantica, Chelophyes contorta, E. mitra y Agalma elegans y algunos physonectes de los géneros Bargmania y Marrus, grupos que aún no han sido suficientemente estudiados en aguas mesopelágicas (Gasca y Suárez-Morales, 1992; Gasca, 1993, 1999a, 1999b; Pugh y Gasca, 2009).
En este trabajo se registran por primera vez a las medusas Halitrephes maasi (Bigelow, 1909); Atorella octogonos Mills, Larson y Young, 1987; Tiburonia granrojo Matsumoto, Raskoff y Lindsay, 2003; Stygiomedusa gigantea (Browne, 1919); Periphyllopsis braweri Vanhöffen, 1902; Crossota millsae Thuesen, 2003; y a los sifonóforos Resomia persicaPugh y Haddock, 2009; R. dunni Pugh y Haddock, 2009; Bargmannia lata Mapstone, 1998 y Lensia quadriculata Pagès, Flood y Youngbluth, 2006, recolectadas u observadas en el golfo de California en expediciones realizadas en 2003 y 2012 por el Monterey Bay Aquarium Research Institute.
A Humberto Bahena por permitirnos usar su fotografía, a Steve Haddock por la identificación de varias de las especies, incluso algunas de las registradas por primera vez en este trabajo. Al MBARI por permitirnos usar la fotografía de Crossota millsae recolectada en aguas del golfo de California.