











Pomacentridae is one of the most abundant fish families in coral and rocky reefs of tropical and temperate waters. Stegastes rectifraenum (Gill, 1862), as other members of the genus, is territorial and actively farm algae on its territory, helping in the regulation of the algal community structure and the reef productivity. It is endemic and one of the most abundant species along the Mexican Pacific coast. Even with its strong relationship with algae, this species is omnivorous. In this analysis, we present the first complete anatomical description of a species of Pomacentridae, focusing in one research question: which structures allow S. rectifraenum to have an omnivorous diet? Through the analysis of the skeletal, muscular and digestive systems, the anatomy of the species was described. The complex anatomy of S. rectifraenum shows a set of characters previously described for grazers (mandibular muscles), herbivorous (mandibular teeth), omnivorous with preference for algae (general skull osteology) and zooplanktivorous species (intestine shape and pharyngeal teeth). According to our results, S. rectifraenum is a grazer, omnivorous with preference for algae. Finally, the key character that facilitates S. rectifraenum access to a wider range of resources is the shape of the pharyngeal teeth, which can effectively grind, fasten and tear.
Pomacentridae es una de las familias de peces más abundantes en arrecifes de coral y rocosos de aguas tropicales y templadas. Stegastes rectifraenum (Gill, 1862), como los miembros del genero Stegastes, es territorial y activamente cultiva algas en su territorio, ayudando en la regulación de la estructura de la comunidad algal y productividad del arrecife. Es endémica y una de las especies más abundantes de la costa del Pacífico mexicano. Aun con su fuerte relación con las comunidades algales, esta especie es omnívora. En este análisis presentamos la primera descripción anatómica completa de un pomacéntrido, enfocándonos en una pregunta: ¿qué estructuras le permiten a S. rectifraenum acceder a una dieta omnívora? A través del análisis de los sistemas óseo, muscular y digestivo, la anatomía de la especie fue descrita. La compleja anatomía de S. rectifraenum muestra un conjunto de caracteres previamente descrito para especies ramoneadoras (músculos mandibulares), herbívoras (dientes mandibulares), omnívoras con preferencia por alga (osteología cefálica general) y zooplanctívoras (forma del intestino y dientes faríngeos). De acuerdo con nuestros resultados, S. rectifraenum es ramoneadora, omnívora con preferencia por algas. Finalmente, el carácter clave que le facilita a S. rectifraenum el acceso a un amplio rango de recursos es la forma de los dientes faríngeos, los cuales pueden moler, sujetar y desgarrar eficazmente.
The family Pomacentridae (damselfishes) is a group of marine fishes found in tropical and temperate waters. It is one of the most abundant fish families in rocky substrates and coral reefs. Worldwide, it groups 394 species (Eschmeyer, 2015). In the Eastern Pacific 7 genera and 24 species have been reported, all endemic to this region (Robertson & Allen, 2006). Damselfishes show an extraordinary diversity of habitat, feeding and behavior preferences (Aguilar-Medrano, Frédérich, De Luna, & E. Balart, 2011). Most of them exhibit aterritorial behavior during the reproductive season, others as the members of the genus Stegastes are always territorial. The species of Stegastes are well known for actively farming and removing unwanted algae growing in its territory (Aguilar-Medrano et al., 2011; Horn, 1989; Wilson & Bellwood, 1997).
In the Tropical Eastern Pacific (TEP), 8 Stegates species have been registered (Robertson & Allen, 2006). Stegastes rectifraenum (Gill, 1862) is endemic to the Mexican Pacific coast and is one of the most widely distributed species in the Gulf of California. It is distributed mostly in rocky reefs, at depths between 1 and 10m, from north of Magdalena Bay on the west coast of Baja California, Gulf of California, Revillagigedo Islands to Acapulco, Guerrero in the south (Allen & Woods, 1980; Robertson & Allen, 2006).
This highly territorial species only leaves its territory for a few seconds to scare intruders or to catch some food. It was first described as a herbivorous species (Robertson & Allen, 2006), feeding mainly on fleshy red and green algae (Montgomery, 1980). However, in spite of its strong relation with algae, new analyses classified this species as omnivorous (Aguilar-Medrano et al., 2011; Moreno-Sánchez, Abitia-Cárdenas, Escobar-Sánchez, & Palacios-Salgado, 2011; Robertson & Allen, 2006).
Stegastes rectifraenum is an important subject of study since it is locally abundant and widely distributed in the Mexican Pacific coast, representing a high contribution to the biomass of shallow coral reefs. Also, it presents highly specialized feeding habits that determine the type of alga that grow in the reef, regulating the algal community structure and thus the productivity of reefs (Montgomery, 1980).
Most osteological analyses of damselfishes have focused just in the skull due to its relationship to diet (Ciardelli, 1967; Emery, 1973, 1980; Frédérich, Parmentier, & Vanderwalle, 2006; Gluckmann & Vanderwalle, 1998; Gluckmann, Bussers, Poulicek, & Vandewalle, 1999; Liem, 1980). Thus, since no complete anatomical description of any damselfish has yet been published, in this study, the anatomical arrangement of S. rectifraenum was described through the combination of the skeletal, muscular and digestive systems. Special attention is given to previously cited structures that are related to the diet in damselfishes, to target our main research question: which structures allow S. rectifraenum to have an omnivorous diet?
Analyses of other members of Pomacentridae, covering different approaches as external morphology, anatomy, gut content, musculature, digestive tract, were used to discuss our results and present a comparative analysis.
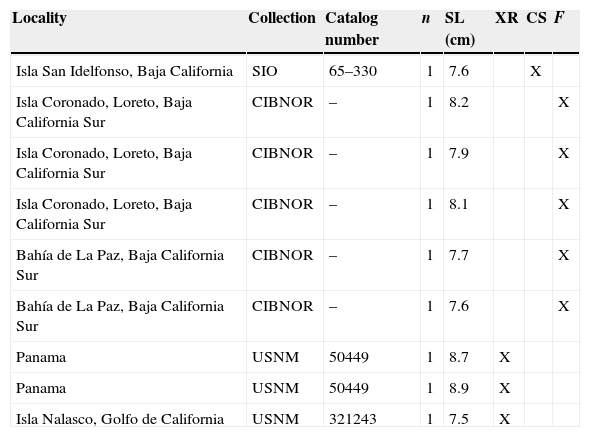
Materials and methodsThe sample comprised 9 adult organisms, from 3 museum collections (Table 1). The analyses of the bone structures, muscle and intestine were carried out by direct observations, observations in stereoscopic microscope and electronic microscope photographs. The osteology was analyzed with 3 techniques: (1) X-rays, (2) fleshing of fresh collected organisms, and (3) clearing and staining technique (Hollister, 1932).
List of the biological material revised from museum collection: (1) Colección de Peces del Centro de Investigaciones Biológicas del Noroeste (CIBNOR), La Paz, B.C.S., Mexico; (2) Collection of marine vertebrates, Scripps Institution of Oceanography (SIO), San Diego, California, USA; and (3) Fish collection of the National Museum of Natural History, Smithsonian, (USNM), Washington, Columbia District, USA. XR: X-ray; CS: clearing and staining technique; F: fleshing.
| Locality | Collection | Catalog number | n | SL (cm) | XR | CS | F |
|---|---|---|---|---|---|---|---|
| Isla San Idelfonso, Baja California | SIO | 65–330 | 1 | 7.6 | X | ||
| Isla Coronado, Loreto, Baja California Sur | CIBNOR | – | 1 | 8.2 | X | ||
| Isla Coronado, Loreto, Baja California Sur | CIBNOR | – | 1 | 7.9 | X | ||
| Isla Coronado, Loreto, Baja California Sur | CIBNOR | – | 1 | 8.1 | X | ||
| Bahía de La Paz, Baja California Sur | CIBNOR | – | 1 | 7.7 | X | ||
| Bahía de La Paz, Baja California Sur | CIBNOR | – | 1 | 7.6 | X | ||
| Panama | USNM | 50449 | 1 | 8.7 | X | ||
| Panama | USNM | 50449 | 1 | 8.9 | X | ||
| Isla Nalasco, Golfo de California | USNM | 321243 | 1 | 7.5 | X |
The terminology used to describe bone structures was based on the analyses of Emery (1973, 1980), Frédérich et al. (2006), Gluckmann and Vanderwalle (1998), Gluckmann et al. (1999), and Kobelkowsky (2004). Ciardelli (1967), Galis and Snelderwaard (1997), Liem (1980), and Winterbottom (1974), were used to describe muscles, and finally, Canan, Silva do Nascimento, Bezerra da Silva, and Chellappa (2012), Fukusho (1969), Kobelkowsky (2002, 2007), and Kobelkowsky & Pantoja-Escobar (2009) were used to describe the digestive system.
ResultsDescription of the skeletal patternNeurocraniumThe neurocranium of S. rectifraenum is slightly longer than higher, the supraoccipital crest is high and rounded (Fig. 1a, b), representing approximately half of the neurocranium height. The inferior extreme of the basioccipital, the anterior extreme of the vomer and the superior extreme of the supraoccipital crest form an angle of approximately 50° (Fig. 1a). The pterotic crest shows a horizontal line of fosses. The posterior edge of the basioccipital coupled with the first vertebrae. The exoccipital and basioccipital mark the dome. In the otic region a wide postemporal fosse is delimited by the pterotic, parietal and epiotic. The parasphenoid form ventrally 2 crests; the posterior one is thinner than the anterior one (Fig. 1d). The vomer is robust and projecting toward the front (Fig. 1c, d); the nasal bones are long and curve and do not exceed the vomer (Fig. 1c). The anterior and superior orbital region has fosses. The suborbital series group 5 bones; the third is the biggest, the edges are serrated (Fig. 1e).
Oral jaws and opercular series left lateral view, (b) posterior view, (c) dorsal view, (d) ventral view, and (e) suborbital bones.")
The upper and lower jaws are short and robust. The premaxilla is robust with narrow and truncated ascending process (Fig. 2a, b, c); the angle between ascending process of the premaxilla and its dentigerous process is equal or less than 90°. The dentigerous area has 1 row of long, sharp-edged teeth whose size gradually decreases from the symphysis (Fig. 2d). The maxilla is robust and undulated with 2 processes in the frontal edge, in between of which the maxilla superior edge is located, the posterior edge is rounded (Fig. 2a, b, c); the lateral surface of the anterior edge is noticeably concave and receives the anterior edge of the palatine (Fig. 2a). The dentary is robust and does not form a coronoid process (Fig. 2a, b, c); at its rear portion presents a process where the articulo-angular joint; presents a row of teeth similar to the premaxilla teeth. The articulo-angular is short (Fig. 2a, b, c), has ample coronoid process and develops a triangular forward process, which is embedded in the dentary; near its posterior edge is the articular fossa, facing upwardly receiving the condyle of the quadrate. The retroarticular is small and joins the articular rear (Fig. 2a, b, c).
 left side view of the mandibular arch, palatine series, jaw suspension, and opercular series; (b) left side view of the upper and lower jaws; (c) medial view of the upper and lower jaws; (d) internal view of the dentary teeth; (e) ventral view of the hyoid apparatus; and (f) left side view of the hyiod apparatus.")
Mandibular skeleton and hyoid apparatus: (a) left side view of the mandibular arch, palatine series, jaw suspension, and opercular series; (b) left side view of the upper and lower jaws; (c) medial view of the upper and lower jaws; (d) internal view of the dentary teeth; (e) ventral view of the hyoid apparatus; and (f) left side view of the hyiod apparatus.
The palatine series is formed by the palatine, ectopterygoid and mesopterygoid bones (Fig. 2a). The palatine is robust and curved; its anterior edge is wider and joins the maxilla, the rear edge articulates with the ectopterygoid and mesopterygoid. The mesopterygoid is short and laminar, and form a fossa that receives part of the adductor arcus palatini muscle. The ectopterygoid is robust and inverted C-shaped; it joins the mesopterygoid and quadrate.
Mandibular suspensionThe mandibular suspension is formed by hyomandibular, metapterygoid, symplectic and quadrate (Fig. 2a). The hyomandibular is elongated and articulates through the condyles with the sphenoid, pterotic and opercular; on its lateral surface forms a curved lip, which join with the upper part of the preopercular; its free anterior edge is slightly convex. The metapterygoid is elongated and joins widely to the hyomandibular. The symplectic is small and oval; near its junction with the quadrate forms a foramen. The quadrate is relatively large and triangular; in its lower-anterior part bear the condyle that articulates with the articulo-angular; its junction with the ectopterygoid is wide; overlap inferiorly with the preopercular.
Opercular seriesIt consists of the preopercular, opercular, subopercular and interopercular bones (Fig. 2a). The preopercular is elongate, L-shaped, with a wide middle portion and convex leading edge; the anterior edge is thicker than the posterior edge; laterally has the lateral line canal; the posterior edge is serrated, mainly the inferior part of the edge. The opercular is trapezoidal, with an upper edge relatively straight and the posterior edge noticeably concave, while its anterior edge is thick and convex. The interopercular is oval and thin, with a wavy upper edge. The subopercular presents thick and rounded edges; has a triangular portion, concave anterior edge and a triangular anterior upper apophysis.
Hyoid apparatusThe hyoid apparatus is relatively small and consists of the basihyal and urohyal odd bones, the 2 hyoid bars which in turn consist of the dorsal hipohyal, ventral hipohyal, ceratohyal, epihyal, interhyal and 6 branchiostegal rays (Fig. 2e, f). The 2 hyioid bars join by the disk-shaped interhyal cartilage, which is a type of diarthrosis (Figs. 3e, 4a).
 left side view of the branchial apparatus and its relation with the basihyal and urohyal; (b) external gill raker of the first branchial arch; (c) external gill raker of the second branchial arch; (d) dorsal view of the lower pharyngeal jaw; (e) ventral view of the upper pharyngeal jaw; (f) dorsal view of the branchial apparatus; (g) ventral view of the branchial apparatus; and (h) frontal view of the lower and upper pharyngeal jaws.")
Branchial apparatus: (a) left side view of the branchial apparatus and its relation with the basihyal and urohyal; (b) external gill raker of the first branchial arch; (c) external gill raker of the second branchial arch; (d) dorsal view of the lower pharyngeal jaw; (e) ventral view of the upper pharyngeal jaw; (f) dorsal view of the branchial apparatus; (g) ventral view of the branchial apparatus; and (h) frontal view of the lower and upper pharyngeal jaws.
 superior view of the lower pharyngeal; (b) lateral view of the lower pharyngeal; (c) superior view of the upper pharyngeal; (d) lateral view of the upper pharyngeal.")
The basihyal is flat shaped in its dorsal view and joins by a diarthrosis to the basibranchial 1 (Fig. 2e). The urohyal is laminar (Fig. 2e) with triangular appearance (Figs. 3f, 4a); its anterior edge is thick and convex, the posterior is concave and the upper shows a condylar structure; at its anterior edge forms 2 lateral processes.
The elements of each hyoid bar are relatively short (Fig. 2e, f). The hipohyales are short. The ceratohyal supports the 6 branchiostegal rays and strongly join the epihyal. The epihyal is rounded and dorsally supports the interhyal, which is small and pyramidal. The branchiostegal rays are 6 elements which gradually increasing in length backwards. The more anterior positioned branchiostegal rays are shorter and thinner, while the others are flat and relatively wide (Fig. 2f).
BranchiocraniumThe first pharyngobranchial keeps the branchial apparatus suspended in the otic region (Fig. 3d). The ceratobranchials are longest than the epibranchials, externally present a groove for the branchial arteries and internally present 2 rows of gill rakers (Fig. 3a, e). The 11–12 gill rakers of the first branchial arch are long and setiform, with 2 series of small and thing teeth (Fig. 3b); the internal row of the branchial arch and 2 rows from the second to the fourth present small gill rakers with setiform teeth in the tip (Fig. 3c). The hyobranchials are 3 thick bones; the second is rectangular; the ceratobranchials and basibranchials fuse to these bones (Fig. 3a, d). The basibranchials are 3 bones, the 2 first are thin and the third is thick (Fig. 3d, e). The glosohyal in thin in the base and it gradually thickens toward the center resembling a shovel.
The lower pharyngeal jaw is elongated, Y-shaped; the anterior edge is thin and toothless, most pharyngeal teeth are in the mesial position (Figs. 3f, 4a, b). Mesial teeth are robust and unicuspid, the teeth close to the edges of the plate are thin, with 1 cuspid resembling a hook with a flat base (Fig. 4a, b). The upper pharyngeal jaw is composed by 4 bones, the 2 largest are thick semi-elliptical plates (Figs. 3g, 4c, d), it presents 2 types of teeth, thick unicuspid teeth in the mesial, and thinner and hook shape teeth in the edges. The 2 smaller plates are triangular with only thin, hook shape teeth (Fig. 4c, d).
Axial skeletonThe vertebral column presents: 3 predorsal bones, 11 precaudal vertebrae, 14 caudal vertebrae and the urostyle. Nine pleural ribs are present from the third vertebra. It presents 27 dorsal pterygiophores, the first 12 join the 12 dorsal fin spines and the next 15 join the 16 dorsal fin rays. The last 2 rays of the dorsal fin are spliced in the base, the last one show a thin base which lay on the base of the penultimate, both rays laid on 15th pterygiophores. The 15th pterygiophore is reduced in size and U-shaped. There are 3 supraneural bones between the supraoccipital crest and the first dorsal pterygiophore. The anal fin is composed by 14 pterygiophores, the first receive the 2 anal spines and the next 13 receive the 14 rays of the anal fin. As in the dorsal fin, the last 2 rays laid in the 13th pterygiophore which is reduced in size and U shaped (Fig. 5a). The caudal complex present 3 epurals, 5 hypurals; the parhypural is thick in the base, tapering to the tip; the ural centrum is thick and the uroneurals 1 and 2 show a long and angular tip (Fig. 5b). The caudal fin presents 25 rays.
Appendicular skeleton vertebral column and support the unpaired fins; (b) caudal complex; (c) pelvic girdle.")
The postemporal join the pectoral girdle to the neurocranium. The 4 radials and the condyle of the scapula support the pectoral fin. The supracleitrum is triangular, laminar, elongated, with the interior edge thick; articulates to the postemporal and cleithrum. The cleithrum is curve with an angular, laminar superior edge and rounded, thick inferior edge. The superior postcleithrum is laminar, thick and elongated. The inferior postcleithrum is laminar, elongated with thick posterior edge. The scapula is small with a central foramen. The coracoid is thin and laminar; the inferior edge joins the inferior edge of the cleithrum, but do not exceed it. The size of the radials increases from the first to the fourth; are thinner in the center, the posterior edge is thicker than the anterior which join the scapula. The radials articulate with 19–21 pectoral rays. The pelvic girdle presents a triangular, thick and angular basipterygium, which join to 2 spines and 5 pelvic rays (Fig. 5c).
Visceral musculatureMandibular musculatureThe more complex muscle of the mandible is the adductor mandibulae, which is divided in the A1, A2, A3 and Aw sections (Fig. 6b, c). Its origin is in several areas of the mandibular suspension and preopercular, and its insertion is in the dentary and premaxilla. The levator arcus palatini muscle is long, originating from a lateral sphenotic area and inserted into the lateral surface of the hyomandibular (Fig. 6b, c, d). The adductor hyomandibulae (not shown) is the antagonist muscle. The adductor arcus palatini muscle originates from the parasphenoid and is widely inserted into endopterygoid, metapterygoid, and part of the hyomandibular (Fig. 6b, c, d). The intermandibularis muscle is narrow and joins the inner surface of both dentary and articular bones, near the mandibular symphysis. It is dorsal and ventral covered by protractor hyoideus (Fig. 7).
 ventral view of the gular muscles; (b) left side view of the adductor mandibulae muscle; (c) the adductor mandibulae with the sections A1 and A2 proyected; and (d) left side view of the palatal muscles.")

The left and right protractor hyoideus muscles originate from the corresponding ceratohyal, above the third branchiostegal ray (Fig. 6a). Both muscles merge in its anterior portion and are attached to the internal part of the dentary, covering the mandibular symphysis. The dilatator operculi muscle is robust, it originates from the lateral surface of sphenotic and inserted in the anterodorsal process of the opercular (Fig. 6b, c). It is largely covered by the levator arcus palatini. The anterior levator operculi muscle (Fig. 6b, c) originates in the pterotic and it is inserted in the front surface portion, near the top edge of the opercular. The levator operculi posterior (Fig. 6b, c) originates in the postemporal and inserted in the upper edge of the operculum, in a posterior position than the anterior levator operculi. The antagonist muscle to these 2 muscles is the adductor operculi (not shown).
Gill musculatureThe externi levatores muscle originates in the sphenotic and is attached in the dorsal process of the 4 epibranchial ridge (Fig. 6). More internally are located the levatores interni (not shown). The retractor dorsalis muscle originates in the lateral surface of the first vertebra and is attached to the posterior edge of the pharingobranchial 3 (Fig. 7). The rectus communis muscle originates in the lateral process of the lower pharyngeal jaws and is inserted into the upper edge of the urohyal (Fig. 6).
The sternohyoideus muscle is strong (Fig. 6), it originates from the anterior surface of the inferior portion of cleitrum and is inserted into the lateral and posterior surfaces of the urohyal. The pharyngoclavicularis externus muscle is laminar; it originates in the middle anterior part of the cleitrum and it is inserted into the ventral surface of the lower pharyngeal bone (Fig. 6). The pharyngoclavicularis internus muscle is laminar, it originates in a higher position of the medium-anterior surface of the cleitrum and is inserted into the ventral surface of the lower pharyngeal bone (Fig. 6).
Visceral cavityThe visceral cavity is bounded by the precaudal vertebrae, the first hemal arch, first anal pterygiophore, the infracarinalis medius muscles, the pelvic girdle, interior infracarinalis muscles, the scapular girdle, the pleural ribs and axial muscles. Approximately one third of the volume of the visceral cavity is occupied by the mesonephric kidneys and gas bladder. The rest of the cavity is occupied by the digestive tract, liver, spleen and gonads. The kidney shapes 2 lateral lobes, from which the left contact the liver. The gaseous bladder contacts the dorsal part of the stomach and gonads. The gonads develop between the gas bladder and bowel, reaching highest volume the mature ovaries, than the mature testicles (Fig. 8a).
Digestive channel left side view of the visceral cavity after removing the body wall; (b) right side view of the digestive tract and liver; (c) left side view of the digestive tract; and (d) ventral view of the liver.")
Immediately behind the upper and lower pharyngeal plates the short esophagus starts. The stomach shows 2 cardiac portions, pyloric and stomach caecum, which are medium sized (Fig. 9a-d). Three pyloric caeca are developed, oriented backwards. One caecum is dorsolateral to the stomach, while the remainders are ventral (Fig. 9d). The intestine is of medium length. Its curl involves the formation of some arcs, after which its trajectory is straight and of ventral position (Fig. 8a, b, c). The anus is bordered with numerous lobules (Fig. 9e) and is partially covered by scales in females (Fig. 9e, f) and totally in males (Fig. 9g). Immediately behind the anus is the urogenital papilla in both sexes.
 dorsal view of the digestive tract and liver; (b) dorsal view of the esophagus, stomach, and pyloric caeca; (c) ventral view of the stomach and pyloric caeca; (d) left side view of the stomach and pyloric caeca; (e) ventral view of the female perianal region; (f) sagittal section of the rectus and urogenital papilla of the female; and (g) ventral view of the male perianal region.")
Digestive tract: (a) dorsal view of the digestive tract and liver; (b) dorsal view of the esophagus, stomach, and pyloric caeca; (c) ventral view of the stomach and pyloric caeca; (d) left side view of the stomach and pyloric caeca; (e) ventral view of the female perianal region; (f) sagittal section of the rectus and urogenital papilla of the female; and (g) ventral view of the male perianal region.
The liver show 2 short anterior lobes, 2 long posterior lobes and some recesses (Fig. 8d). The left posterior hepatic lobe is the broader and partially covers the stomach and pyloric caeca (Fig. 8a). The right hepatic lobe develops lesser extent (Fig. 8b, d). The gallbladder is elongated and slender, and is attached to the posterior edge of the right hepatic lobe (Fig. 9a).
DiscussionOur analysis is the first presenting a complete description of the skeletal, muscle and digestive systems of a damselfish species. Although, damselfishes appear similar in their external morphology (Allen, 1991), our detailed anatomical examination and the analysis of the literature allow us to observe differences among S. rectifraenum and other damselfish species. According to our revision, the key to have a wider diet is related to a peculiar anatomical pattern, combining structures observed in herbivorous, omnivorous and zooplanktivorous species.
The cephalic region of S. rectifraenum shows diagnostic characters of the genus, as serrate margins of the preopercular and suborbital bones. The cranium is high and compact, the pieces are in general thick and the supraoccipital crest is high and rounded. In contrast, Lepidozygus tapeinosoma (Bleeker, 1856) shows a low and long cranium, the pieces are thin and the supraoccipital crest is low and small (Emery, 1980). These morphological differences are related to the general shape pattern, which is more hydrodynamic in L. tapeinosoma than in S. rectifraenum.
Is interesting to notice that the urohyal bone of S. rectifraenum, with its triangular appearance, is highly similar to that of Microspathodon chrysurus (Cuvier, 1838) (Ciardelli, 1967; Emery, 1973), and different from L. tapeinosoma (Emery, 1980), and some Chromis species, which present elongated urohyal bones (Emery, 1973). This character, as well as the general shape of the cranium, might be related to the general shape of the species; while Stegates and Microspathodon are deep-bodied species, the members of Lepidozygus and some Chromis species are elongated.
Stegates rectifraenum presents 3 predorsal bones as most damselfish species (Emery, 1980). The number, shape and relation of the dorsal fin and neural spines are generally consistent within species but diverge between species, genera or families (Kendall, 1976). For example, in S. rectifraenum the first predorsal is wider, while the other 2 are thinner and all its superior-lateral processes are forward inclined. In contrast, L. tapeinosoma predorsal bones are about as wide, just the first and third present superior-lateral processes and each is inclined in opposite direction (Emery, 1980).
Another character that shows differences among species is the caudal complex. The caudal complex has been used to help in the classification of fish groups and provide useful information about swimming (Collette & Russo, 1984; Laidig & Adams, 1991; Schultze & Arratia, 1989). Among this complex, the presence of 5 hypurals has been recognized as a character of advanced teleosts, however in some fish groups as the mackerel and tunas the hypurals fuse to form a hypural plate, while in others as the scombrids, hypurals 1–2 and 3–4 are united (Helfman, Collette, Facey, & Bowen, 2009). Damselfishes might be highly variable in this character, since S. rectifraenum present 5 hypurals, non-united and L. tapeinosoma present the hypurals 1–2 and 3–4 united (Emery, 1980).
Trophic tendenciesStegastes rectifrenum presents 1 row of premaxillar teeth, a character related to herbivorous species as M. chrysurus (Emery, 1973; Frédérich et al., 2006; Gluckmann & Vanderwalle, 1998) and the mandibular arch has thick and compact pieces, character related to grazer damselfish species as Plectroglyphidodon lacrymatus (Quoy & Gaimard, 1825) (Frédérich et al., 2006).
Also, it presents characters related to omnivorous species with preference for algae as some Dascyllus, Chrysiptera and Pomacentrus species: the dentary is almost flat and the retroarticular is long and thin; the premaxillar and dentary present incisor teeth; the teeth of the dentary are shorter than the premaxillar teeth; the anterior teeth are longer than the laterals; the angle of the ascending process of the premaxilla and the dentate area form an angle 90° approximately; and the connection between the metapterigoides and quadrate is thin (Frédérich et al., 2006; Frédérich, Pilet, Parmentier, & Vanderwalle, 2008; Gluckmann & Vanderwalle, 1998; Norman & Greenwood, 1963).
The levator arcus palatine is long in S. rectifraenum and short in M. chrysurus (Ciardelli, 1967) both are approximately the same width. The main function of this muscle is to raise the hyomandibular, participating in complex movements of swallowing (Emery, 1973). Strong, thick mandibular muscles are a character related to herbivorous (Vial & Ojeda, 1990), thus, this character in S. rectifraenum might be related to an omnivorous feeding behavior.
Most of the internal organs in S. rectifraenum are similar to M. chrysurus, as the left lobe of the liver is longest, the gall bladder is located dorsomedially in the right lobe of the liver; 3 pyloric caeca, etc. (Ciardelli, 1967), however the intestine of M. chrysurus occupied more space in the visceral cavity than S. rectifraenum intestine. The size of the intestine is a character strongly attributable to diet; generally speaking, herbivorous species have long intestines (Canan, Silva do Nascimento, Bezerra da Silva, & Chellappa, 2012; Elliott & Bellwood, 2003; Emery, 1973; Emery & Thresher, 1980; Horn, 1989). It is clear that S. rectifraenum presents shortest intestine than M. chrysurus, and also, the intestinal convolutions of S. rectifraenum show strong relation to zooplanktivorous species as some Chromis, Abudefduf and Chrysiptera species (Fukusho, 1969).
The pharyngeal teeth of S. rectifraenum are similar to those in pelagic feeders as some Chromis and Abudefduf species (Emery, 1973). These highly efficient teeth, can effectively grind, fasten and tear, and might be the key character that allows S. rectifraenum to use a wider range of resources, avoiding or decreasing competition when a specific resource is limited in the environment. Finally, through the complete anatomical analyses we classify S. rectifraenum as grazer, omnivorous, with preference for algae.
Ecological implicationsAlso when the consumption of algae of S. rectifraenum just represents half of its diet (Aguilar-Medrano et al., 2011; Moreno-Sánchez et al., 2011; Robertson & Allen, 2006), this species spends the full day aggressively protecting the territory from intruders (Aguilar-Medrano, Frédérich, Balart, & De Luna, 2013; Ceccarelli, 2007; Lobel, 1980). Thus, the algae in S. rectifraenum territory might play different roles as: (1) food (Aguilar-Medrano et al., 2011; Moreno-Sánchez et al., 2011; Robertson & Allen, 2006); (2) a substrate that accumulates debris, which is part of its diet (Aguilar-Medrano et al., 2011; Wilson & Bellwood, 1997); and (3) attracting small benthic invertebrates, since approximately 50% of its diet is composed of invertebrates and other animals. Since S. rectifraenum barely leaves its territory, then the territory must attract the prey.
To the Consejo Nacional de Ciencia y Tecnología, México funded the research of R. Aguilar-Medrano. We are grateful to the fish collections that donated the material, Centro de Investigaciones Biológicas del Noroeste, Scripps Institution of Oceanography, and Smithsonian National Museum of Natural History. The manuscript benefited from comments by two anonymous reviewers and the associate editor Leticia Huidobro.
Peer Review under the responsibility of Universidad Nacional Autónoma de México.