





La estacionalidad ambiental afecta la biología y ecología de las especies de lagartijas, sin embargo, sus respuestas dependerán de las características del hábitat. A la fecha, no hay trabajos que comparen el efecto de la estacionalidad en poblaciones de lagartijas insulares y continentales de una especie. Nuestro trabajo aborda el efecto de la estacionalidad en el uso del hábitat y en la actividad de 2 poblaciones de lagartijas Anolis nebulosus, una insular y una continental en la costa de Jalisco, México. Los resultados indican que las fluctuaciones estacionales en la temperatura ambiental y en la humedad relativa afectan el uso de la percha y la actividad de ambas poblaciones; sin embargo, la población insular presentó mayor actividad y difirió en el uso del hábitat con respecto a la población continental, lo que indica que las condiciones ambientales y ecológicas en la isla son menos agresivas para la población de lagartijas Anolis nebulosus cuando se comparan con las del continente.
Environmental seasonality affects the biology and ecology of lizards, however, they may respond depending of the habitat characteristics. Until now, there are not studies about the effect of seasonality in insular lizard populations and its comparison with mainland populations. In our work we measured seasonality effects upon habitat use and activity of 2 Anolis nebulosus lizard populations, 1 insular and 1 continental, in Jalisco coast, Mexico. Our results suggest that seasonal fluctuations in environmental temperature and relative humidity affect perch use and activity of both populations; however, insular population was more active and differs in habitat use from the continental population, suggesting that environmental and ecological conditions on the island are less aggressive for Anolis nebulosus lizard, when compare with the continental population.
La estacionalidad ambiental que incluye cambios en la precipitación, en la temperatura del aire, disponibilidad de alimento y fotoperiodo (Fleming y Hooker, 1975), tiene un gran impacto en la ecología, biología y comportamiento en las especies de lagartijas (Adolph y Porter, 1996; Fleming y Hooker, 1975; García, Valtierra-Azotla y Lister, 2010; Gienger, Beck, Sabari y Stumbaugh, 2002). Algunas de las estrategias desarrolladas por este grupo para enfrentar los efectos de la estacionalidad incluyen: modificaciones en su ciclo y niveles de actividad, uso del hábitat (Fleming y Hooker, 1975; García et al., 2010; Gienger et al., 2002), tasa de crecimiento (Adolph y Porter, 1996) y condición corporal (Colli, Pérez y Zatz, 1997), entre otras. Sin embargo, el efecto de la estacionalidad se puede manifestar a través de diferentes factores que dependen del hábitat. Por ejemplo, en zonas templadas las lagartijas deben enfrentar fluctuaciones estacionales marcadas en la temperatura ambiental, por lo que la disponibilidad y calidad de los sitios de asoleo o termorregulación son importantes para el desarrollo de sus actividades físicas y sus procesos fisiológicos (Van Damme, Bauwens y Verheyen, 1987), mientras que en las zonas tropicales, la cantidad y frecuencia de la precipitación tienen un gran impacto en la dieta de los lacertilios a través de los cambios en la disponibilidad de alimento (Stamps, 1977) y en su actividad por cambios en la cobertura del dosel (Gienger et al., 2002).
Las islas son hábitats que suelen tener características particulares, debido, principalmente a su extensión y distancia con la costa (Kadmon y Pulliam, 1993). Diversos trabajos en lagartijas insulares indican cambios morfológicos, biológicos (deshidratación, daño corporal y mortalidad) y ecológicos (traspasamiento de nicho, densidad poblacional y competencia) en respuesta a las condiciones de aislamiento (Case y Niche, 1983; Klütsch, Misof, Grosse y Moritz, 2007; Macrini, Irschick y Losos, 2003; Perry, Dmi’el y Lazell, 1999; Vervust, van Dongen, Grbac y van Damme, 2009). Sin embargo, no se ha abordado el posible efecto que tiene la estacionalidad sobre esas poblaciones insulares y si su respuesta es similar a la de sus contrapartes continentales.
En este trabajo se abordará el efecto de la estacionalidad de las lluvias sobre la actividad y comportamiento de la lagartija Anolis nebulosus (A. nebulosus) en una isla y en un área continental, que se encuentran distanciadas por 6.5km, con el fin de contestar las siguientes preguntas: ¿Las variaciones estacionales en la temperatura ambiental y la humedad relativa son similares en los 2 hábitats (continental e insular)? y ¿cómo responde la lagartija A. nebulosus a los cambios estacionales en la isla y en el continente?
Materiales y métodosEl estudio se llevó a cabo dentro de los terrenos de la Estación de Biología Chamela (19°29’48.61” N, 105°02’25.89” O) que forma parte de la Reserva de la Biósfera Chamela-Cuixmala y en la isla San Agustín (19°32’04.63” N, 105°05’18.08” O) que pertenece al Santuario de las Islas de la Bahía de Chamela, ambos sitios localizados en la costa de Jalisco, México. El clima encontrado en ambos lugares pertenece a los cálidos-húmedos con precipitación anual que varía entre 700 y 1,000mm y una temperatura anual promedio de 24.9°C. La característica que resalta en esa región es la estacionalidad de las lluvias, que se concentran de junio a octubre con una temporada de sequía prolongada que va de noviembre a junio (Ceballos y Miranda, 2000). La vegetación que predomina en ambos sitios de estudio es el bosque tropical caducifolio, aunque el encontrado en la isla San Agustín es menos desarrollado (ver Ceballos, Szekely, García, Rodríguez y Noguera, 1999; Conanp, 2008; Noguera, Vega, García y Quesada, 2002).
A. nebulosus es una lagartija pequeña (LHC: 35-47mm) que se caracteriza por presentar un abanico grande y desplegable de color naranja con borde blanco, más grande y notorio en machos que en hembras. La coloración del dorso es gris y el vientre es blanco, es una lagartija arborícola, ovípara e insectívora, es endémica de México y se distribuye del este de Sonora hacia el norte de Sinaloa y de ahí por toda la vertiente del Pacífico hasta el sur de Guerrero, además, en la cuenca del río Balsas (García y Ceballos, 1994; Ramírez-Bautista, 1994).
El trabajo de campo se realizó durante las estaciones seca (abril-mayo) y lluviosa (septiembre-octubre) de 2008 y se dividió en 2 partes. En la primera parte se trazaron 3 transectos de 100×5m en cada sitio, los cuales fueron recorridos de manera alterna entre la isla y el continente durante 8 semanas de cada temporada de trabajo. Asimismo, en cada lugar se alternó la hora de visita o recorrido entre 3 horarios: a) 09:00-11:00 am; b) 11:00 am-01:00 pm y c) 01:00-03:00 pm, horario aproximado de actividad diurno de la especie de estudio (ver Lister y García, 1992). De esta manera, al final de cada temporada de trabajo se realizaron 54 recorridos para cada transecto por estación, en cada sitio de estudio. Durante cada recorrido se hizo un registro de todos los individuos de A. nebulosus encontrados, y de cada uno se determinó el sexo, con base en la presencia o ausencia de abanico gular, así como la altura de la percha en la cual se observó. Además, cada 10min o al observar una lagartija, se registró la temperatura ambiental y la humedad relativa, factores para medir la estacionalidad, usando para ello un termohigrómetro IDT® precisión ±1°C y ±5%, respectivamente.
En la segunda parte una persona realizó observaciones focales de individuos adultos de A. nebulosus, siguiéndolos el mayor tiempo posible hasta perder contacto visual, en cuyo caso se buscó un nuevo individuo; antes de iniciar las observaciones se dio un tiempo de 5min para permitir que el individuo se acostumbrara a la presencia del observador (Lister y García, 1992). Para evitar alterar el comportamiento de los individuos, el seguimiento se hizo a una distancia superior a 3m, algunas veces se utilizaron binoculares. Para cada individuo observado se registró el sexo, se promedió el número de movimientos realizados por hora, la distancia recorrida por movimiento y el tiempo transcurrido entre movimientos. Asimismo, se promedió la altura, diámetro y distancia de la percha a la cual atacan a sus presas, y movimiento donde el individuo presentó actividad masticatoria. Finalmente, se registró la frecuencia de movimientos en diferentes horarios del día y la frecuencia de movimientos dedicados a las siguientes actividades: a) termorregulación: que se refiere a cambios de postura y desplazamientos sin fin de alimentación o socialización; b) socialización: movimientos seguidos por despliegues que indican interacción con otros individuos (ver Jenssen, 1971) y c) alimentación: que se refiere a movimientos en los que los individuos presentaron actividad masticatoria.
Análisis estadísticoLos datos de las variables de comportamiento medidas no se ajustaron a una distribución normal (Kolmogorov-Smirnov, p>0.05), por lo que se usó un análisis Kruskal-Wallis para contrastar dichas variables entre sitios, estaciones y sexos. La prueba Kruskal-Wallis (K-W test) es frecuentemente usada en trabajos de comportamiento, ya que está diseñada para analizar datos con escala de medición débil (Wayne, 2012). Los datos de temperatura ambiental, humedad relativa y frecuencia de movimientos a lo largo del día (3 horarios: a) 8-11h, b) 11-14h y c) 14-17h) fueron procesados por sitio y estación; mientras que los datos de altura de la percha, movimientos por hora, distancia por movimiento, distancia por hora, tiempo entre movimientos y altura, diámetro y distancia de ataque a las presas se procesaron por sitio, estación y sexo. Los datos de tipo de actividad se analizaron solo para machos, debido a que el comportamiento suele medirse más en machos que en hembras (Melville, 2002) y a que fueron pocos los datos que se registraron en las hembras. Todos los datos de actividad fueron analizados por individuo.
ResultadosEstacionalidad ambientalLa temperatura ambiental (K-W test=45.02, df=7, p<0.0001) y la humedad relativa (K-W test=426.06, df=3, p<0.001) presentaron diferencias significativas entre sitios; sin embargo, la temperatura en ambos lugares fue similar durante la estación seca, pero mayor en la isla que en el continente durante la estación lluviosa (fig. 1A). La humedad relativa fue similar en ambos sitios durante la estación lluviosa, pero mayor en la isla durante la estación seca (fig. 1B). Se obtuvo información de la altura de la percha utilizada por 602 individuos en total durante los recorridos en los transectos, que corresponden al uso de percha de 29 hembras continentales, 82 machos continentales, 70 hembras insulares y 421 machos insulares. Se registraron diferencias estadísticamente significativas en la altura de la percha usada entre sitios y entre estaciones (K-W test=55.53, df=7, p<0.001), los machos percharon a mayor altura que las hembras en ambos sitios, mientras que las hembras y los machos insulares percharon a mayor altura que sus contrapartes continentales. En ambos sitios, las hembras percharon a mayor altura durante la estación seca (fig. 2).
 y continente (C) durante las estaciones seca (S) y lluviosa (Ll). Intervalo de confianza del 95%. Se muestra que entre grupos hay diferencia significativa (Dif. Sig.).")
 y hembras (H) de la isla (I) y continente (C) durante las estaciones seca (S) y lluviosa (Ll). Intervalo de confianza del 95%. Se muestra que entre grupos hay diferencia significativa (Dif. Sig.).")
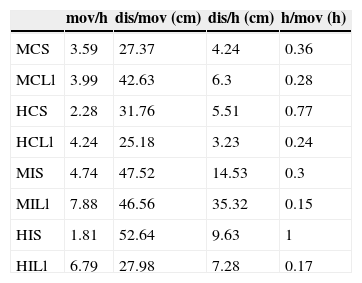
Se lograron observaciones focales de 61 individuos, 34 en la isla y 27 en el continente, que totalizaron 348h de observación, 161h en la isla y 186h en el continente. El tiempo promedio de observación por individuo fue de 4.8h en la isla y de 6.91h en el continente. Ya que los datos obtenidos se analizaron por individuo, los análisis que se presentan a continuación se realizaron con un tamaño de muestra de 61, a menos que se indique lo contrario. La comparación entre machos de ambos sitios registró diferencias significativas en la frecuencia de movimientos por hora (K-W test=24.11, df=7, p=0.001), en la distancia total recorrida por movimiento (K-W test=14.19, df=7, p=0.047), en la distancia total recorrida por hora (K-W test=24.11, df=7, p=0.001) y en el tiempo que transcurre entre movimientos (K-W test=27.035, df=7, p=0.0003). En promedio, las lagartijas insulares se movieron más frecuentemente, recorrieron mayor distancia por movimiento y se desplazaron más que las continentales (tabla 1). En los machos insulares se registró mayor frecuencia de movimientos por hora, distancia por movimiento y distancia recorrida por hora, que en los continentales (tabla 1). Durante la estación lluviosa se registraron los valores más altos de movimiento por hora (tabla 1). El tiempo entre movimientos indica que los machos de la isla y durante la estación lluviosa son más activos (tabla 1). Las hembras presentaron similitud en su actividad tanto entre sitios como entre estaciones, excepto en los movimientos por hora que fueron mayores en la estación lluviosa y en la isla (tabla 1).
Promedio de movimientos por hora, distancia por movimiento, distancia por hora y tiempo entre movimientos de machos y hembras en la isla y el continente durante la estación seca y la estación lluviosa.
| mov/h | dis/mov (cm) | dis/h (cm) | h/mov (h) | |
|---|---|---|---|---|
| MCS | 3.59 | 27.37 | 4.24 | 0.36 |
| MCLl | 3.99 | 42.63 | 6.3 | 0.28 |
| HCS | 2.28 | 31.76 | 5.51 | 0.77 |
| HCLl | 4.24 | 25.18 | 3.23 | 0.24 |
| MIS | 4.74 | 47.52 | 14.53 | 0.3 |
| MILl | 7.88 | 46.56 | 35.32 | 0.15 |
| HIS | 1.81 | 52.64 | 9.63 | 1 |
| HILl | 6.79 | 27.98 | 7.28 | 0.17 |
C: continente; dis/h: distancia por hora; dis/mov: distancia por movimiento; h/mov: tiempo entre movimientos; H: hembras; I: isla; Ll: estación lluviosa; M: machos; mov/h: promedio de movimientos por hora; S: estación seca.
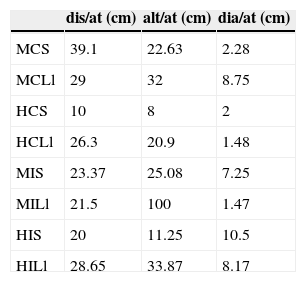
La distancia (K-W test=3.73, df=7, p=0.809), altura (K-W test=6.73, df=7, p=0.457) y diámetro (K-W test=9.04, df=7, p=0.249) de la percha en la cual atacan a sus presas, no presentaron diferencias significativas entre sitios. La distancia de ataque a las presas de A. nebulosus varía, entre estaciones, más en el continente que en la isla. De la estación seca a la lluviosa los machos disminuyeron la distancia de ataque y las hembras la incrementaron (tabla 2). La altura de la percha en la cual atacan a sus presas es mayor en las lagartijas insulares que en las continentales y mayor en los machos que en las hembras en ambos sitios (tabla 2). El diámetro de la percha en el cual atacan a sus presas tiende a disminuir en la estación lluviosa, excepto para los machos continentales (tabla 2).
Promedio de la distancia de ataque, altura de la percha de ataque y diámetro de la percha de ataque de machos y hembras en la isla y el continente durante la estación seca y la estación lluviosa.
| dis/at (cm) | alt/at (cm) | dia/at (cm) | |
|---|---|---|---|
| MCS | 39.1 | 22.63 | 2.28 |
| MCLl | 29 | 32 | 8.75 |
| HCS | 10 | 8 | 2 |
| HCLl | 26.3 | 20.9 | 1.48 |
| MIS | 23.37 | 25.08 | 7.25 |
| MILl | 21.5 | 100 | 1.47 |
| HIS | 20 | 11.25 | 10.5 |
| HILl | 28.65 | 33.87 | 8.17 |
C: continente; alt/at: altura de la percha de ataque; dia/at: diámetro de la percha de ataque; dis/at: promedio de la distancia de ataque; H: hembras; I: isla; Ll: estación lluviosa; M: machos; S: estación seca.
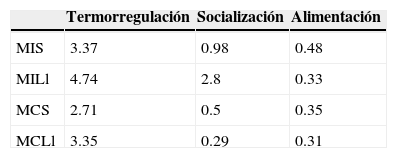
No se registraron diferencias significativas en los movimientos dedicados a termorregular (K-W test=5.3, df=3, p=0.15), socializar (K-W test=5.85, df=3, p=0.11) o alimentarse (K-W test=0.52, df=3, p=0.91) entre ambos sitios (tabla 3). La frecuencia de movimientos que realizan a lo largo del día no presentó diferencias significativas (K-W test=12.91, df=7, p=0.074) entre sitios. Machos y hembras continentales presentaron actividad bimodal durante la estación seca, pero no durante la estación lluviosa (fig. 3), mientras que machos y hembras insulares disminuyeron su actividad conforme avanzó el día durante la estación seca, pero no durante la estación lluviosa (fig. 3).
Promedio de movimientos por individuo para termorregular, socializar y alimentación de machos en la isla y el continente durante la estación seca y la estación lluviosa.
| Termorregulación | Socialización | Alimentación | |
|---|---|---|---|
| MIS | 3.37 | 0.98 | 0.48 |
| MILl | 4.74 | 2.8 | 0.33 |
| MCS | 2.71 | 0.5 | 0.35 |
| MCLl | 3.35 | 0.29 | 0.31 |
C: continente; H: hembras; I: isla; Ll: estación lluviosa; M: machos; S: estación seca.
 8-11h, 2) 11-14h y 3) 14-17h en machos (M) y hembras (H) de la isla (I) y el continente (C) durante las estaciones seca (S) y lluviosa (Ll).")
Durante la realización del trabajo se registró una mayor temperatura ambiental y menor humedad relativa durante la estación seca, lo cual se debe a que en ambos sitios la vegetación predominante es el bosque tropical caducifolio, vegetación que pierde la mayoría del follaje durante dicha estación, permitiendo la entrada de los rayos del sol (Gienger et al., 2002) y el aumento de la temperatura bajo el dosel. Sin embargo, también se observó una menor fluctuación de ambas variables en la isla San Agustín. Esto puede deberse a que en la isla, a diferencia del continente, el viento sopla gran parte del año (Conanp, 2008), levando humedad (Hill y Lee, 1957) y temperatura a su interior (Bovo, Marques y Andrade, 2012) y consecuentemente, podría amortiguar las altas temperaturas y bajos porcentajes de humedad relativa.
Un trabajo previo indica que A. nebulosus responde a la estacionalidad en las lluvias bajando su actividad y disminuyendo la altura a la cual percha (Lister y García, 1992). Los resultados de nuestro estudio con relación a la altura de la percha indican que los machos continentales y las hembras de ambos sitios disminuyeron la altura a la cual perchan durante la estación lluviosa. La disminución de la percha que se registró en las hembras del presente estudio puede permitir la división de recursos con los machos y, consecuentemente, evitar o disminuir la competencia con ellos (Zucker, 1986). En cuanto a los machos continentales, es probable que con la disminución de la altura a la cual perchan eviten la competencia con Sceloporus melanorhinus que es una lagartija arborícola de mayor tamaño y que suele perchar a grandes alturas (García et al., 2010). Sin embargo, cabe la posibilidad de que la mayor cobertura vegetal en el continente con respecto a la isla San Agustín (Conanp, 2008) haya dificultado la observación de individuos en perchas altas durante la estación lluviosa.
La disminución en la actividad durante la estación seca concuerda con lo observado en Anolis cupreus (Fleming y Hooker, 1975), Anolis carolinensis (Ballinger, 1977), Anolis aeneus (Stamps y Tanaka, 1981), Sceloporus melanorhinus y Sceloporus utiformis (García et al., 2010), incluyendo lo observado por Lister y García (1992), sin embargo, en contraste con este último trabajo, la diferencia que encontramos en la actividad entre la estación seca y la estación lluviosa no es tan marcada. La diferencia entre ambos trabajos probablemente se debe a los tiempos de muestreo; Lister y García (1992) trabajaron en los meses de febrero y julio de 1987 y en marzo y julio de 1988 cuando los individuos se alimentan y cortejan con fines reproductivos, mientras que el trabajo de campo en este estudio se ubicó en los meses de abril-mayo y en septiembre-octubre, cuyos periodos están más lejos de la actividad reproductora máxima (Ramírez-Bautista y Vitt, 1997). Durante la máxima actividad los machos son más activos, ya que su objetivo es defender un territorio que les permita acceso a las hembras con fines reproductivos, mientras que las hembras son más activas para incrementar sus reservas de energía, las cuales se traducirán en producción de huevos (Andrews, 1971; Stamps, 1982).
ActividadLos datos sobre la actividad a lo largo del día (fig. 3) indican cambios estacionales, muy similares a los encontrados por Lister y García (1992) con actividad bimodal en la estación seca y actividad variable en la estación lluviosa en el continente. La actividad bimodal permite a las lagartijas del género Anolis evitar las temperaturas altas registradas a mediodía en la estación seca, cuando no hay cobertura del dosel y así evitar su deshidratación (Heatwole y Verón, 1977) y permitir la acumulación de energía (Colli et al., 1997; Floyd y Jenssen, 1983). En la estación lluviosa la cobertura vegetal las cubre de los rayos directos del sol y de esta manera pueden permanecer activas. Por su parte, las lagartijas insulares no presentan actividad bimodal durante la estación seca y parece que la mayor parte de su actividad la llevan a cabo durante la mañana y mediodía, ya que es probable que las corrientes de aire ya mencionadas permitan su actividad durante esas horas. No obstante, la actividad tanto de machos como de hembras se distribuye de manera más homogénea durante la estación lluviosa, que sería similar a lo que sucede con la población continental en esa estación.
InsularidadLos resultados de este trabajo indican que los individuos de la isla San Agustín perchan a mayor altura y son más activos que los encontrados en el continente. El ambiente menos fluctuante en la isla puede facilitar el mejor desempeño en las lagartijas insulares, mientras que las lagartijas A. nebulosus continentales podrían gastar más energía para mantener una temperatura corporal adecuada, perjudicando su actividad y condición corporal, como se ha observado en otras lagartijas (Herczeg, Gonda, Saarikivi y Merilä, 2006).
Al comparar nuestros resultados con los registrados por Lister y García (1992), encontramos mayor actividad de las lagartijas A. nebulosus en el continente y aún mayor en la isla tanto en la estación seca como en la estación lluviosa. La actividad reducida de A. nebulosus en la región de Chamela se ha atribuido a la estacionalidad ambiental y a la depredación (Lister y García, 1992). La depredación y la competencia interespecífica se consideran interacciones que tienen una influencia importante en la ecología y evolución en este grupo de lagartijas (Calsbeek y Cox, 2010, 2011). Sin embargo, se considera que en las islas estas 2 interacciones pierden importancia (Buckley y Roughgarden, 2006), aumentando, generalmente la densidad poblacional y consecuentemente la competencia intraespecífica (Thomas, Meiri y Phillimore, 2009). La competencia intraespecífica podría estar explicando los resultados sobre actividad, ya que los individuos necesitan interactuar más para la defensa de su territorio; asimismo puede explicar los resultados de uso de la percha, ya que en los machos y hembras insulares se marca más la diferencia en la altura a la cual perchan que en el continente, indicando con ello mayor competencia entre ellos (Zucker, 1986). La estacionalidad ambiental afecta la actividad y uso del hábitat de poblaciones insulares y continentales de lagartijas A. nebulosus, sin embargo, condiciones ambientales menos fluctuantes en la isla que en el continente permiten mayor actividad en perchas con mejores cualidades para patrullar el territorio de las lagartijas insulares, condiciones que también podrían facilitar la disminución de la depredación y la competencia interespecífica.
Al Posgrado en Ciencias Biológicas de la Universidad Nacional Autónoma de México por la beca otorgada.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.