


El objetivo de la presente revisión es demostrar que, en actividades breves y de alta intensidad, el uso exclusivo del consumo de oxígeno subestima significativamente el cálculo del gasto energético total, y que la medición de la acumulación del lactato en sangre es una alternativa válida en tales casos.
Actualmente no existe una metodología ideal para cuantificar el gasto energético que no es generado por el metabolismo aeróbico. La biopsia muscular es un método directo; no obstante es invasivo y ha de aceptarse que una minúscula muestra refleja los eventos metabólicos del músculo en su totalidad. Por otro lado, el EPOC (exceso de consumo de oxígeno posejercicio) no mide el calor liberado en la transformación del piruvato a lactato (proceso irreversible).
Margaria et al., demuestran que la tasa de acumulación de lactato en sangre (en g/kg de peso corporal/minuto) aumenta de manera lineal con la potencia metabólica (en kcal/kg de peso corporal/minuto) de un trabajo. Di Prampero propone calcular la ecuación de regresión de la pendiente de la línea que describe dicha relación, a la cual denomina como β, coeficiente que da paso a la cantidad de energía por unidad de masa corporal aportado por la acumulación de 1 mMol de lactato en sangre y su valor es 3,0 ml O2/kg de peso/mMol.
En actividades breves e intensas, la utilización del [Δ lactato sanguíneo] como indicador del gasto energético no aeróbico es una herramienta sumamente valiosa, a pesar de no ser totalmente exacta.
The aim of this review is to show that exclusively using oxygen consumption for brief and high intensity activities considerably underestimates total energy expenditure calculation. Alternatively, measuring lactate accumulation in blood appears to be usefulness for such purpose.
Currently, there is no an ideal methodology to quantify to the non-aerobic energy expenditure. Muscle biopsy is a direct but invasive method. Moreover, it involves assuming that a very small sample reflects whole muscle´s metabolic events. On the other hand, the excess post-exercise oxygen consumption does not measure the heat released through the conversion of piruvate into lactate (irreversible process). Margaria et al., show that the rate at which lactate is accumulated (expressed as g/kg of body weight / minute) increases linearly with the corresponding metabolic power (in kcal/kg of body weight/minute). Di Prampero proposes to calculate the reciprocal of the slope which describes this relationship (which he calls β). Such a coefficient means the amount of energy per unit of body weight supplied by the accumulation of 1mMol of lactate in blood and its value is 3.0 ml O2/kg of weight/mMol.
During brief and intense activities, the fact of using the [Δ blood lactate] as a measurement of the non-aerobic lactic energy expenditure is a highly valuable tool, even though it is not completely precise.
Introducción
La estimación del gasto energético de distintos tipos de actividades físicas y en diferentes circunstancias es de suma importancia en ámbitos diversos como pueden ser el ejercicio físico, la salud y el trabajo. El análisis del gasto energético producido en diferentes tipos y niveles de esfuerzo ha servido especialmente para el desarrollo de tablas de referencia habiendo sido estudiado tanto en poblaciones adultas1-7, como en niños y adolescentes8 y también en preescolares9.
Desde los años 70, ya existen estudios interesados en la estimación del gasto metabólico centrado específicamente en esfuerzos de corta duración. Así, destaca el trabajo pionero de Wilmore et al.10 en 1978, en el que estudian el gasto metabólico experimentado con tres circuitos de diez ejercicios de fuerza, hallando una media de gasto calórico de 9,0 kcal/min para los hombres y 6,1 kcal/min para las mujeres. Más adelante, Hempel y Wells11, analizan un programa Nautilus Express (programa de ejercicios de fuerza con periodos de descanso breves) de 20 minutos y reportan un gasto calórico promedio de 5,13 kcal/min para las mujeres y 7,80 kcal/min para los hombres. El interés por la comparación del gasto calórico entre hombres y mujeres en esfuerzos de corta duración ha continuado, y trabajos más recientes analizan entre otras respuestas, el gasto calórico en un circuito (6 ejercicios de 12 repeticiones) de entrenamiento con peso libre12, o en circuitos de entrenamiento de fuerza con máquinas con un amplio rango de intensidades13. Farinatti et al14, estiman el gasto energético para demostrar la existencia de diferencias cuando se alteran las secuencias de ejercicios de un circuito de pesas. En otros trabajos se evalúa el gasto calórico en entrenamientos de fuerza en circuitos para validar metodologías como monitores metabólicos portátiles15, o simplemente en diversas poblaciones como estudiantes universitarios16 o en cardiópatas17, incluso comparando el gasto energético de circuitos de pesas con carrera en tapiz18. Otras publicaciones se centran en estimar el gasto calórico de un solo tipo de ejercicio19-21, o entrenamientos basados en series durante varios días22,23.
Uno de los motivos quizás más polémicos por el que se ha estimado el gasto energético durante esfuerzos de corta duración, ha sido el de analizar la validez de los entrenamientos de fuerza para satisfacer las demandas de gasto energético propuestas en las recomendaciones del Colegio Americano de Medicina del Deporte (ACSM) para mantener un adecuado nivel de salud y aptitud cardiorrespiratoria24,25. En esta línea, Becham y Earnest26 analizan el entrenamiento con peso libre considerando que genera mayor gasto metabólico por solicitar éstos el trabajo de musculatura estabilizadora, pero demuestran que no provee un estímulo de entrenamiento cardiovascular suficiente. Otro estudio27 se centra en ejercicios de resistencia funcional continua (CFE), en el que obtienen valores de gasto energético, no de consumo de oxígeno, que alcanzaron los requeridos en las recomendaciones del ACSM. Por último, Phillips y Ziuriatis28 en 2003, concluyen que el protocolo de entrenamiento de fuerza del ACSM24,25 requeriría un mayor número de series, ejercicios o repeticiones, para poder alcanzar el volumen absoluto mínimo de 150 kcal reportado por el propio ACSM como necesario para alcanzar beneficios de salud que se alcanzan con ejercicios de endurance.
Cabe mencionar, que para llegar a sus conclusiones, los autores del trabajo anteriormente mencionados28 realizan el cálculo del gasto calórico midiendo el consumo de oxígeno durante el esfuerzo y luego transformando éste en 5,05 kcal por litro de oxígeno consumido. De la misma manera, se ha empleado el consumo de oxígeno para la realización de las diferentes tablas de gasto calórico de distintos tipos de actividades, lo que ha llevado a datos discutibles y de dudosa validez en algunos casos.
La calorimetría indirecta, calculada a través de la medición del consumo de oxígeno, es un método simple y su uso está sumamente extendido como estimador de la producción de calor. No obstante, en esfuerzos físicos muy intensos en los que existe un importante aporte energético que no proviene del metabolismo oxidativo, la medición del consumo de oxígeno como único método para calcular el gasto calórico total, puede llevar a subestimar dicho valor (más aún si sólo se tiene en cuenta el consumo de oxígeno intraesfuerzo)29.
En el presente trabajo de revisión se pretende remarcar que cuando la rápida resíntesis glucolítica de ATP (adenosín trifosfato) excede al ritmo del trabajo mitocondrial con la consiguiente producción de lactato y de calor, la medición del consumo de oxígeno como única metodología no refleja con precisión el gasto energético total que se lleva a cabo, y esto hace necesario el empleo de una metodología que estime el gasto calórico que no proviene del metabolismo aeróbico como es el uso de la acumulación del lactato, método que no está exento de imperfecciones, pero que puede ser útil, al menos en actividades breves y de alta intensidad.
Bioenergética: algunos conceptos a tener en cuenta
Flujo de energía, ineficiencia energética: calor y entropía
Independientemente del motivo por el que se valora el gasto energético de una actividad física, la mayor parte del mismo proviene del generado durante la contracción muscular. Los elementos que intercambian energía durante la contracción muscular y el metabolismo se acoplan, ya que por un lado existe una conversión y transferencia de energía químicoquímica a través de las vías metabólicas (desde los enlaces químicos de los nutrientes de los alimentos a los enlaces químicos de las moléculas de ATP) y, por otro lado, sucede una conversión y transferencia químicomecánica a través de la contracción muscular (desde los enlaces químicos del ATP al trabajo muscular). En definitiva, la energía de la hidrólisis del ATP en el músculo debe ser suficientemente importante como para promover el movimiento de las fibras musculares. Cabe mencionar, no obstante, que aunque no se suelen tener en cuenta, existen otros tipos de gastos energéticos como son los generados por las diferentes bombas de iones, la biosíntesis de carbohidratos, grasas, y proteínas, señales intra y extracelulares, etc.
El flujo de energía que sucede en el interior celular se realiza de manera descendente, esto es de una situación de menor entropía (energía que no está disponible para realizar trabajo) a uno de mayor entropía. Dicho esto de manera más sencilla, la transferencia de energía es ineficiente y esta ineficiencia siempre aparece en forma de producción de calor que usualmente es desechado al ambiente30.
En un sistema abierto, la energía libre de Gibbs (energía disponible para realizar trabajo) puede cambiar según varíe el ritmo de la transferencia de energía y la proporción producto/reactante durante el intercambio (por ejemplo conforme se altera la distancia del equilibrio)31. En el interior de las células, la entropía y la producción de calor son el resultado continuo de la transferencia de energía durante la hidrólisis del ATP y su regeneración, conocidos ambos procesos de manera conjunta como resíntesis de ATP. Mientras que la entropía no puede medirse directamente, la pérdida de calor se puede medir por calorimetría directa o ser estimada indirectamente por el consumo de oxígeno32.
El intercambio de gases y su relación con el gasto de energía
A finales del siglo XIX, investigadores como Pfluger, realizan comparaciones entre medidas directas de producción de calor con mediciones indirectas de intercambio de gases. Como consecuencia de estos estudios, en los que analizaron la combustión de compuestos orgánicos, se halló que existía una alta correlación lineal entre consumo de oxígeno y la entalpía (contenido calórico de un sistema, reflejo del número y tipo de enlaces químicos en los reactivos y los productos). Posteriormente, se demostró que la relación entre el calor producido y el oxígeno consumido era similar para la combustión de compuestos orgánicos y la respiración celular, -110 kcal por mol de O2, ya que la producción de entalpía por equivalente de electrón es aproximadamente de -27,5 kcal por mol de O2 (un átomo de C tiene cuatro valencias)30.
Cuando lo que se oxida es una dieta compuesta en su totalidad por carbohidratos, el RER (tasa de intercambio respiratorio, relación CO2/O2) es 1,0 (6 CO2/ 6 O2), y se considera que la producción de calor es 5,02 kcal por litro de oxígeno consumido (1l de O2 = 5,02 kcal). Cuando los ácidos grasos son los principales substratos oxidados, el RER es 0,7 (la oxidación del aceite de palma = 16 CO2/ 23 O2) y se considera que el consumo de 1l de O2 produce 4,68 kcal (1l de O2 = 4,68 kcal) de calor33. Si se tienen en cuenta los valores del RER, se podría interpretar que los carbohidratos son una fuente más eficiente de combustible por usar menos oxígeno que las grasas durante su oxidación, aunque lo que sucede en realidad, es que el piruvato para transformarse en acetil coA y entrar al ciclo de Krebs debe descarboxilarse (pérdida de 1 CO2 con acción de la piruvato deshidrogenasa PDH), mientras que las grasas se van dividiendo en la beta-oxidación mitocondrial en moléculas de dos carbonos directamente. Por volumen de ATP resintetizado, la oxidación completa de la glucosa si la comparamos con la oxidación de las grasas tiene una producción relativa de CO2 adicional, y no un menor consumo de oxígeno relativo.
La producción de calor y el intercambio de gases, sean estos productos de la respiración celular o de la combustión de compuestos orgánicos, se relacionan de igual manera. La mayor producción de calor (0,34 kcal) por equivalente de oxígeno que se produce cuando se oxidan carbohidratos respecto de las grasas (5,02 kcal frente a 4,68 kcal), podría ser atribuida a la producción de transferencia de energía en forma de calor y entropía durante la glucólisis hasta la formación de piruvato (2 de los 36 ATP producidos en la glucólisis se obtienen antes de que se ingrese al ciclo de Krebs)33.
Metabolismo aeróbico y no aeróbico
Cuando se habla de metabolismo no aeróbico, se está haciendo referencia a aquella situación en la que por diversas causas (ritmo de la glucólisis, tasa de aceptación por parte de la mitocondria del piruvato formado), parte de la obtención de energía para la realización del trabajo celular se lleva a cabo sin la utilización de oxígeno. Esto no significa necesariamente la ausencia de oxígeno en el interior celular durante la glucólisis rápida, ni que dicha circunstancia esté generando este tipo de metabolismo, al menos no es la única causa, ni lo más habitual durante el esfuerzo.
La resíntesis aeróbica y la resíntesis no aeróbica de ATP pueden ser entendidas, desde un punto de vista metabólico, como procesos independientes, ya que cada una tiene diferentes reactantes y productos, usan diferentes enzimas y diferentes tipos de reacciones químicas, se dan en compartimentos celulares distintos, explotan diferentes tipos de gradientes, y cada uno funciona con diferente eficiencia34. En aquellas circunstancias en las que la tasa de fosforilación glucolítica (con 2 ATP; 0,36 kcal por l de O2) es igual a la tasa de respiración mitocondrial (con 36 ATP; 4,68 kcal por l de O2) ambos componentes metabólicos, el aeróbico y el no aeróbico de la glucólisis pueden sumarse e interpretar una resíntesis conjunta de 36 ATP y una conversión del gasto de energía de 5,04 kcal por litro de O2 consumido.
Producción y aclaramiento de lactato
La contracción muscular, al igual que el resto de las actividades celulares, requiere de energía que es aportada por el ATP. Por lo tanto, en el interior de las células que están trabajando existe una continua hidrólisis de ATP en ADP y Pi, y, dada la escasa cantidad del primero, su resíntesis debe ser continua. La glucólisis es uno de los procesos encargados de aportar la energía necesaria para volver a formar ATP. En un ejercicio al 60 % del VO2máx., como resultado de la degradación de una molécula de glucosa se liberará energía suficiente para regenerar dos moléculas de ATP y se terminarán formando dos moléculas de piruvato (de 3 carbonos cada una), NADH y protones que serán luego consumidos como sustrato de la respiración mitocondrial (cuando el ritmo de la glucólisis es igual al de la res piración mitocondrial). Cuando el ejercicio es de gran intensidad, la hidrólisis de ATP se realiza a un ritmo tal que su resíntesis no puede ser sostenida al 100% por la respiración mitocondrial, ya que el ritmo de la glucólisis supera al ritmo de trabajo de la mitocondria. Por lo tanto, la acumulación de piruvato y NADH generados necesita de un aceptor de esos protones, y de esta manera se forma lactato (a pH fisiológico no se produce ácido láctico, conversión del 99% a lactato)35-37 siendo el NADH oxidado a NAD.
Durante los ejercicios de corta duración y de alta intensidad, las fibras glucolíticas del músculo aumentan la producción de lactato y su aclaramiento disminuye, resultando en un incremento de su concentración intramuscular. Esto genera que aumente su liberación a la sangre, aunque una parte puede difundir a fibras musculares oxidativas vecinas donde luego puede ser oxidado. El lactato liberado a la sangre puede ser captado por los músculos que están en reposo, o que trabajan a intensidades bajas o moderadas. Por lo tanto, la mayoría del lactato aclarado de la sangre se emplea como combustible oxidativo, dependiendo su tasa absoluta tanto de los músculos que trabajan como de los que están en reposo38-43, una parte del lactato sanguíneo es consumido como combustible por el corazón y otra es usada para la gluconeogénesis por el hígado, entre otros destinos.
Aunque se haya mencionado que la relación entre la producción de calor y el consumo de oxígeno durante la respiración es de -110 kcal por mol de O2, en el trabajo de Gnaiger y Kemp44 realizado con cultivo de células de mamífero, el valor de esta relación varió de -117 a -191 kcal por mol de O2 o más. Para los autores de la investigación, esta mayor cantidad de calor que va de -7 a -81 kcal por mol de O2 consumido, se debería a un incremento en la formación de lactato en los cultivos de células de mamíferos, y a un presumible aumento de la contribución no aeróbica en el gasto de energía a la resíntesis total de ATP; es decir, que la aceleración en la producción de lactato contribuye sustancialmente a la producción de calor más allá de la participación mitocondrial. Si el calor sirve como medida estándar para la medición del gasto metabólico, tanto la glucólisis como la glucogenólisis rápidas con la consiguiente producción de lactato poseen el potencial de hacer significativas contribuciones al gasto energético celular30. Pasteur demostró que la utilización de la glucosa en las levaduras era más rápida cuando el oxígeno estaba ausente45. Una vez más, se debería considerar que la medición exclusiva del consumo de oxígeno no refleja con precisión el ritmo de resíntesis de ATP que se lleva a cabo en estas circunstancias.
Algunos autores señalan, que además de aportar ATP, la fosforilación glucolítica rápida sirve para: mantener el potencial redox46, proteger las células contra el estrés oxidativo47, promover la formación de precursores biosintéticos del crecimiento celular48 y como mecanismo de control de crecimiento celular. Independientemente de las funciones que cumpla, la formación de lactato está asociada a producción de calor y entropía, y por definición, ineficiencia y gasto de energía. Debería tenerse en cuenta que el paso más importante para la producción de calor durante la glucólisis es la reducción de piruvato a lactato49 (-15 a -191 kcal por mol de O2) siendo irreversible este gasto de energía en forma de calor, como se argumentará más adelante.
Gaesser y Brooks50 describen los diferentes destinos de la remoción del lactato y del piruvato, los cuales son: convertirse en glucosa en el hígado (ciclo de Cori) en glucógeno en las células (glucógeno-génesis) o en alanina. El lactato, además de su oxidación completa, también puede ser aclarado a través de la oxidación aeróbica completa del piruvato, y argumentan de esta manera que la "deuda de oxígeno" no representa adecuadamente el gasto energético glucolítico anaeróbico. Es más, en un sistema abierto, ambas reacciones bioquímicas, la aeróbica y la no aeróbica, a menudo suceden lejos de estar en equilibrio51.
Según Di Prampero52, cuando la producción de lactato se iguala con su aclaramiento, permaneciendo constante por lo tanto su concentración en los fluidos corporales (lo que sucede en ejercicios suaves/aeróbicos), la tasa de consumo de oxígeno es un reflejo global del gasto de energía de todo el cuerpo, sin importar la magnitud de la producción y aclaramiento de lactato o la concentración absoluta de lactato. Por el contrario, en ejercicios intensos, donde la concentración de lactato continúa aumentando, el intercambio de energía en todo el cuerpo es mayor que el consumo de oxígeno en una cantidad que es proporcional a la tasa de acumulación neta de lactato.
Medición del gasto energético no aeróbico
Actualmente, no existe un método ideal para cuantificar el gasto energético no aeróbico y las metodologías propuestas como gold standard son poco adecuadas en actividades intensas y de corta duración, además de ser costosas y poco prácticas53. En este contexto, el gasto de energía suele ser estimado mediante el uso exclusivo del consumo de oxígeno, con el error que esto conlleva subestimando el gasto energético total en aquellas actividades que son sustentadas en gran medida por un metabolismo no aeróbico.
De las diferentes técnicas existentes para la estimación del gasto energético no aeróbico, la biopsia muscular podría ser de utilidad, ya que aporta reflejo directo del intercambio de energía que sucede en el interior celular. No obstante, es un proceso invasivo (con el inconveniente que ello significa) y se debe asumir que una minúscula muestra muscular representa fielmente los eventos metabólicos, aeróbicos y no aeróbicos, de la totalidad del músculo, el cual está constituido de miles de fibras, las que además pueden ser de diferentes tipos en cuanto a sus características metabólicas.
Entre los métodos no invasivos para estimar el gasto energético no aeróbico (ATP-CP y glucólisis rápida), destacan dos de ellos en los cuales uno requiere de la medición de lactato sanguíneo y el otro del déficit de oxígeno. El primero solo estima el aporte de la glucólisis rápida, mientras que la medición del déficit de oxígeno estima el aporte de ambos componentes no aeróbicos del gasto metabólico. No obstante, cabe señalar, que en esta última metodología, en lo que se refiere a los depósitos de fosfágenos, sólo representa el uso del ATP-CP y no su resíntesis, la cual ocurriría en la recuperación, y es por ello que para autores como Sott54 el déficit de oxígeno es de utilidad cuando se quiere medir el gasto energético durante el ejercicio exclusivamente, pero que no lo es si se desea estimar el gasto del ejercicio y de la recuperación.
Las opciones habituales para cuantificar el gasto de energía en un ejercicio de tipo no aeróbico y su recuperación son altamente cuestionables55: por un lado, la suma del déficit de oxígeno más la medición del consumo de oxígeno del ejercicio, en donde se estaría ignorando el gasto de energía durante la recuperación; por otro lado, el consumo de oxígeno del ejercicio más el EPOC (exceso de consumo de oxígeno postejercicio), en cuyo caso se estaría ignorando el metabolismo no aeróbico.
El EPOC se basa en la teoría de que la medición del consumo de oxígeno de la recuperación del ejercicio representa el gasto energético no aeróbico que sucede durante dicha actividad56. Dicho de otra manera, sería una devolución aeróbica durante la recuperación de una deuda no aeróbica contraída durante el esfuerzo. Incluso se consideraba que durante la recuperación existía una primera fase de declinación rápida del consumo de oxígeno que sería aláctica (resíntesis de fosfágenos) y una parte del consumo de oxígeno posesfuerzo debida al aclaramiento oxidativo del lactato57. Gaesser y Brooks50, en su revisión sobre las bases metabólicas del exceso de consumo de oxígeno posejercicio fundamentan y concluyen que el aclaramiento del lactato y el consumo de oxígeno posejercicio no están relacionados ni temporal ni causalmente. Durante la recuperación de ejercicios de alta intensidad puede generarse un elevado gasto energético, y durante la misma se resintetiza ATP-CP, de la misma manera que otros procesos que demandan energía como la restauración de la hemoglobina y la mioglobina, el aumento de la circulación y la ventilación, recambio de proteínas, ciclo de las grasas, resíntesis de glucógeno a partir del lactato57.
Por otro lado, debemos tener en cuenta que el paso de piruvato a lactato es el proceso de mayor producción de calor de la glucólisis49 y que dicho paso, al ser llevado a cabo en un sistema abierto es un proceso de pérdida de calor irreversible54. Scott y Kemp58 en un trabajo llevado a cabo con preparados celulares de fibras cardiacas que respiraban unas con aportes externos de piruvato y otras con aporte de lactato, hallan que la producción de calor de ambos tipos de preparados celulares es equivalente cuando se expresa por mol de oxígeno consumido. Por lo tanto, demuestran que no se consume calor cuando el lactato es reconvertido a piruvato, la reacción no es reversible desde un punto de vista termodinámico, y la transferencia de energía durante la respiración mitocondrial no representa la transferencia de energía en la forma de resíntesis de ATP glucolítica rápida con formación/acumulación de lactato. Los autores concluyen que sus hallazgos indican que la producción de lactato y su oxidación son eventos de gasto de energía separados que necesitan ser sumados para interpretar adecuadamente el gasto de energía total.
Existen otras circunstancias en las que se demuestra que no siempre el consumo de oxígeno refleja realmente la producción total de calor. Davies y Brindle59, en su trabajo con células aisladas (levaduras) manipuladas genéticamente para incrementar el contenido de fosfofructoquinasa (enzima glucolítica que cataliza la transformación de fructosa 6 fosfato a fructosa 1-6 bifosfato), hallaron que la resíntesis no aeróbica de ATP se incrementaba, y esto se acompañaba de un descenso del VO2 del 36%, pero sin que cambiara la resíntesis de ATP respecto a la de las células no manipuladas. Esto puede interpretarse como que, al menos en células aisladas, existiría una diferencia entre el gasto de energía como pérdida de calor y el consumo de oxígeno.
Por lo tanto, es correcto asumir que la producción de ATP durante la transformación pasajera de piruvato a lactato no se ve alterada, pero el calor producido y luego liberado en esta reacción está claramente afectado. Esto hace que sea inconcebible pensar que este calor generado por la glucólisis no aeróbica sea representado por la medición del consumo de oxígeno mitocondrial durante la recuperación. La contracción muscular que utiliza como combustible la glucólisis rápida aumenta de manera irreversible el calor y la entropía del entorno, los cuales no son luego aclarados por la mitocondria durante la oxidación del lactato44. Es decir, el gasto de energía asociado con el ATP de la glucólisis rápida y la producción de lactato que excede al metabolismo aeróbico no puede ser representado más tarde por la medición del consumo de oxígeno (EPOC).
La utilización del [Δ lactato sanguíneo] como indicador del gasto energético no aeróbico
El lactato recibe su nombre porque fue hallado originariamente en la leche. Como producto de la glucólisis no aeróbica, el lactato es una molécula de glucosa que se ha dividido por la mitad. A pesar de ello, mientras que moderados niveles de glucosa son considerados de manera positiva, el lactato, históricamente, ha sido considerado problemático, cualquiera que sea su concentración37 (sea en músculo o en sangre).
Como se detalló con anterioridad, cuando el requerimiento de energía de un ejercicio es muy elevado, una parte del ATP es resintetizado por la glucólisis no aeróbica. En tales circunstancias, y después de los 20-30 segundos iniciales que tarda el VO2 en alcanzar su valor máximo, se podría considerar que el total de la resíntesis del ATP es igual a la cantidad de ATP resintetizada por el VO2máx más la cantidad de ATP resintetizada por la tasa neta de acumulación de lactato producido por los músculos. Esto ha llevado a que algunos autores60-64 hayan utilizado la acumulación de lactato sanguíneo como índice de la glucólisis no aeróbica, ya sea esto para la realización de cálculos teóricos y/o por razones prácticas. No obstante, como la producción neta de lactato no puede ser medida directamente, y su estimación indirecta puede ser considerada poco fiable, otros autores65,66 han negado su utilidad.
Ole Bang67, en 1936, afirma que durante un ejercicio físico prolongado, independientemente de su intensidad o duración, ocurre habitualmente un pico en los niveles de concentración de lactato sanguíneo a los diez minutos de empezado el ejercicio, y luego comienzan a declinar durante el transcurso del mismo. A la luz de este hallazgo, muchos investigadores consideraron que el lactato, a priori, no era fiable como indicador válido del metabolismo glucolítico no aeróbico (gasto energético no aeróbico). Esta afirmación es de aplicación en el caso de ejercicios que duran diez minutos o más, mientras que en aquellos que duren entre 1-10 minutos, donde el lactato tiene siempre su pico de concentración en sangre durante la recuperación del ejercicio, y no en el transcurso del mismo, la realidad es diferente. Por lo tanto, se puede considerar que si bien los niveles de lactato sanguíneo y muscular no son en verdad un medio perfecto de cuantificar la resíntesis de ATP glucolítica no aeróbica, bajo condiciones específicas bien definidas (ejercicios breves de alta a muy alta intensidad, y solamente cuando el lactato tienes su pico en la recuperación y no durante el ejercicio), puede ser de utilidad ayudando a aportar una razonable estimación del gasto energético glucolítico no aeróbico.
En 1963, Margaria et al.68, determinaron el gasto energético que ocurría en dos atletas de media distancia, al correr en tapiz rodante con diferentes velocidades (de 9 a 22 km/h) y distintas inclinaciones (de -20 a +15%), permitiendo de esta manera estimar los correspondientes requerimientos de potencia metabólica. En otro estudio69, determinaron la tasa de acumulación de lactato en sangre a intensidades (velocidades e inclinaciones de tapiz) que requerían una potencia metabólica superior a la del consumo máximo de oxígeno, de dos estudiantes no atletas y un atleta olímpico de media distancia. Cada una de estas velocidades supramáximas fue dividida en varias series (4 o 5) con una duración incremental (2-10 minutos), hasta el agotamiento voluntario, realizándoles extracciones de sangre al final del esfuerzo y durante la recuperación (minutos 1, 2, 5, 8, 15 y 30). De esta manera, pudieron demostrar que la tasa de acumulación de lactato en sangre (en g/kg de peso corporal/minuto) se incrementaba de manera lineal con la potencia metabólica (en kcal/ kg de peso corporal/minuto) del ejercicio. La pendiente de dicha curva fue tomada como medida de la cantidad de energía liberada en vivo por la producción de un gramo de lactato: alcanzaba a 22 cal/gramo de lactato (20 kcal/mol). Este hallazgo original fue confirmado posteriormente por otros estudios llevados a cabo con perros61 y en músculo aislado (gemelo) de perro70. Para el cálculo del lactato "producido" por kilogramo de peso corporal, los autores asumieron que el pico de concentración de lactato en sangre entre el minuto 5-8 de la recuperación era una medición del lactato en equilibrio en todos los fluidos corporales y que esto era igual en todos los tejidos, sin tener en cuenta las diferentes fracciones de agua intra y extracelulares (esto no es del todo así, ya que los aniones de lactato se localizan preferentemente en la fase extracelular). Además, no consideraron que una cierta cantidad de lactato se aclara inevitablemente en los primeros cinco minutos de la recuperación, antes de tener el pico de concentración en sangre, y por lo tanto dicho valor es menor de lo que sería en realidad. No obstante esto no fue motivo para desestimar el método.
Para Di Prampero71, el hecho de que la cinética de la desaparición del lactato de la sangre sea de manera monoexponencial permite asumir:
1) Que la concentración pico de lactato en sangre es el resultado de una condición de equilibrio, la cual es independiente de la concentración existente en los fluidos extra e intracelular.
2) Que las tasas de desaparición de lactato en los diferentes compartimentos de fluidos corporales tienen la misma constante de tiempo. De tal manera, que si se estandarizan las condiciones de la recuperación, el pico de concentración de lactato en sangre durante la recuperación es directamente proporcional a la cantidad total de lactato acumulado en un 1 kg de masa corporal durante el ejercicio precedente supramáximo, y además, la tasa de acumulación de lactato en sangre, calculada ésta a partir del ratio "pico de concentración de lactato/duración del ejercicio" precedente supramáximo, es directamente proporcional a la tasa de acumulación de lactato en 1 kg de masa corporal.
En otras palabras, no hay necesidad de determinar la tasa de acumulación de lactato por kg de masa corporal para obtener información de la energía aportada por el metabolismo no aeróbico láctico. Di Prampero71 propone que sería suficiente con calcular la ecuación de regresión de la pendiente de la línea que describe la medición de la tasa del incremento del lactato en sangre en relación con la potencia metabólica que se requiere para la realización de dicho ejercicio (ya conocida). A este parámetro se lo denominó como "β" y es en realidad el coeficiente que da paso a la cantidad de energía por unidad de masa corporal aportado por la acumulación de 1 mMol de lactato en sangre.
En experimentos posteriores al de Margaria et al.68, y bajo condiciones cuidadosamente definidas, se continuó demostrando la existencia de relaciones lineales entre las concentraciones sanguíneas de lactato y las cargas de trabajo en diferentes tipos de poblaciones y esfuerzos físicos. Por un lado, el propio Margaria et al.72, determinan un valor de β equivalente a 3,0 ml O2/kg de peso/mMol en 12 sujetos de diferentes niveles de aptitud corriendo en tapiz a diferentes intensidades supramáximas. Pendergast et al.73, evaluando a 64 nadadores de competición (42 varones y 22 mujeres) obtienen un valor de β para la natación de 2,7 ml O2/kg de peso/mMol y, finalmente Capelli et al.74, analizando a 11 ciclistas de alto nivel, encuentran valores de β intermedios entre los obtenidos por Margaria et al.72 y los de Pendergast et al.73. Scott54 centró sus análisis en experiencias con levantamiento de peso. El ejercicio elegido fue press de banca, y el trabajo realizado se estimó con la asistencia de un dispositivo electrónico que medía la distancia de levantamiento. Los participantes del estudio realizaron siete series de levantamientos en siete días distintos, a los que se les midió la concentración de lactato en el minuto dos de la recuperación. Los datos del estudio piloto aportados por Scott, una vez más denotan una correlación lineal entre el ejercicio físico realizado y la concentración de lactato en sangre.
Para di Prampero52, la notable similitud de estos valores de β, independiente del tipo de sujetos, tipo e intensidad de los ejercicios y la variada musculatura involucrada, refuerza la visión de que β es en realidad el equivalente energético de la acumulación de lactato sanguíneo, y sustenta su validez general. No debería considerarse β como un equivalente energético de formación de lactato en los músculos activos, y no aporta ninguna información directa sobre la relación estequiométrica entre la formación de lactato y la resíntesis de ATP. No obstante, es una medida que permite determinar la energía liberada en el cuerpo siempre que la concentración de lactato aumenta una determinada cantidad posteriormente a la realización de un ejercicio breve e intenso. En definitiva, lo que se debería tener en cuenta, por lo tanto, es que el significado energético del lactato depende de la tasa a la cual cambia su concentración en los fluidos corporales y no de su concentración absoluta71.
A partir de los datos de los trabajos antes mencionados57,68,72-74, se considera que β tiene un valor de 3,0 ml O2/kg de peso/mMol, lo que significa que cada 1 mMol de aumento de la concentración de lactato en sangre equivale a la energía aportada por el consumo de 3,0 ml O2 por kg de peso corporal.
En definitiva, para estimar el gasto energético proveniente de la glucólisis rápida mediante esta metodología se debe calcular primero [Δ lactato sanguíneo] que se obtiene de medir la concentración del lactato pico (aproximadamente 2 minutos de la recuperación) y restarle la concentración del lactato basal. A la diferencia obtenida, [Δ lactato sanguíneo], se la transforma en equivalente de oxígeno de la siguiente manera:
1 mMol de [Δ lactato sanguíneo] = 3,0 ml de O2/ kg de peso corporal
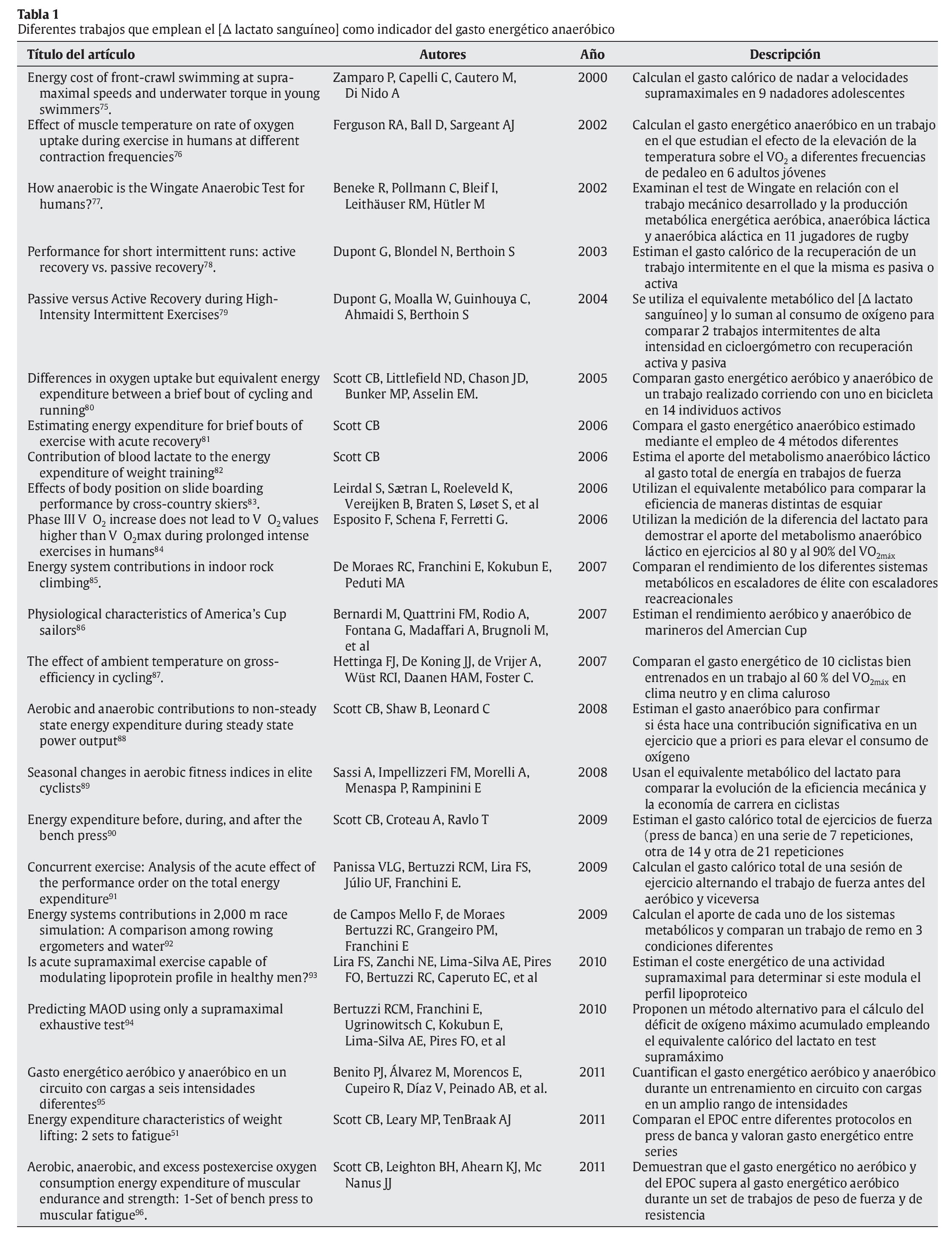
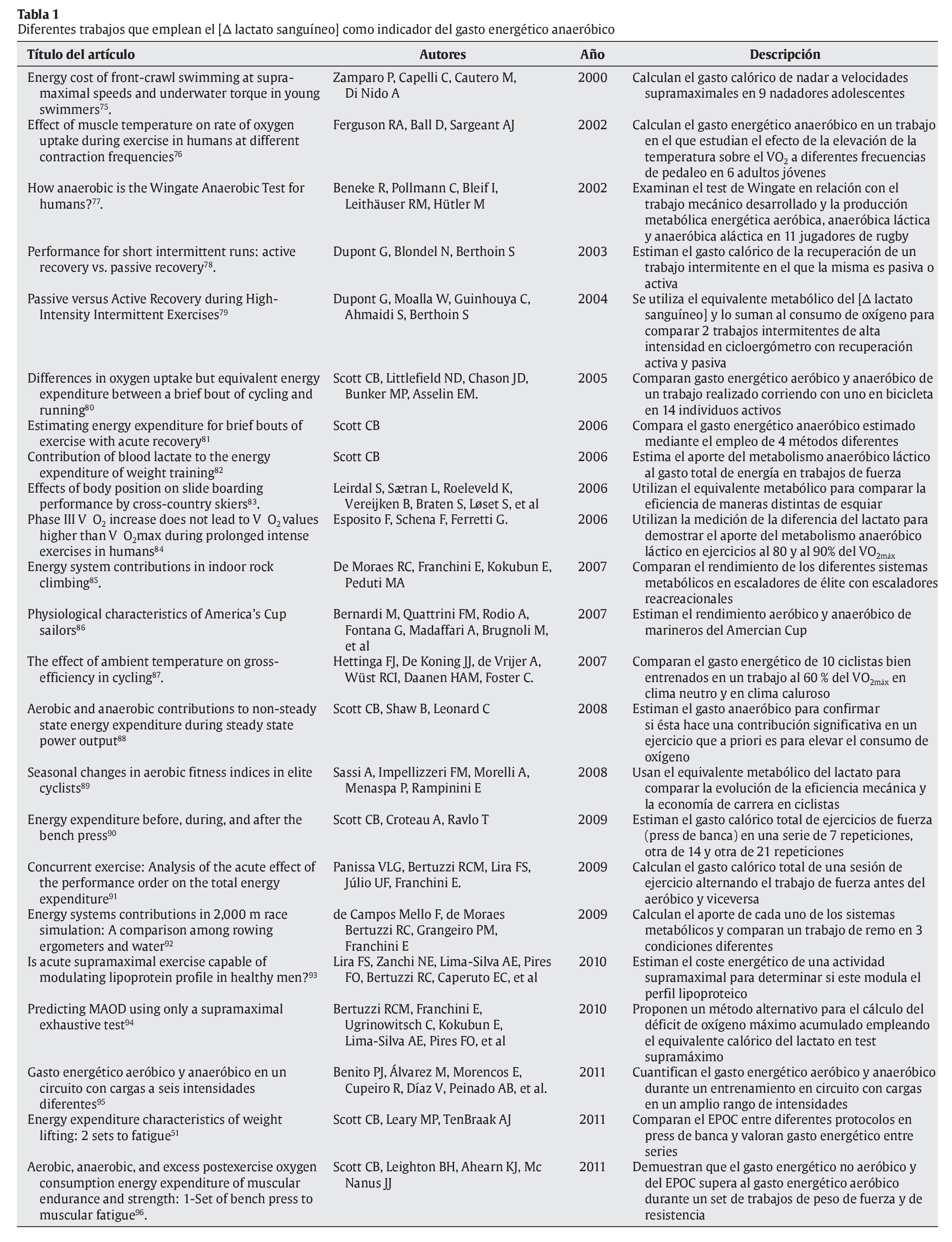
Al igual que cuando se calcula el gasto energético proveniente del metabolismo aeróbico, luego se transforma el equivalente de oxígeno obtenido en 5,05 kcal por litro de oxígeno. Esta propuesta metodológica ha sido utilizada en algunos trabajos con objetivos variados y en la tabla 1 se aportan algunos de ellos.
Conclusiones y aplicaciones prácticas
La estimación del gasto energético en actividades de intensidad leve a moderada y de larga duración es relativamente simple a través de una metodología ampliamente utilizada como es el consumo de oxígeno. No obstante, existe el problema de estimar adecuadamente el gasto metabólico no aeróbico. Existe más que demostrada evidencia de que para ejercicios de alta intensidad y corta duración en los que se produce una elevación de la concentración del lactato sanguíneo durante la recuperación, la medición del ritmo de la acumulación del lactato (equivalente calórico a partir del [Δ lactato sanguíneo]) es una razonable y útil metodología, a pesar de sus limitaciones. La no inclusión de dicha valoración sería perpetuar un error histórico cuando se pretende valorar el gasto total de una actividad física que tenga un importante aporte no aeróbico a dicho gasto.
Desde un punto de vista práctico, para la estimación del gasto calórico total de una actividad física, se deberían medir tanto:
1) El gasto energético no aeróbico glucolítico mediante el equivalente calórico del [Δ lactato sanguíneo]
2) El gasto energético aeróbico del ejercicio mediante el consumo de oxígeno y su posterior transformación en equivalente calórico.
3) El gasto energético aeróbico (no glucolítico) de la recuperación y su posterior transformación en equivalente calórico.
Correspondencia:
Dr. B. Hernán Viana-Montaner.
Centro Andaluz de Medicina del Deporte.
Plaza de Vistalegre, s/n. Puerta Oeste.
14004 - Córdoba. España.
Correo electrónico: bernardoh.viana@juntadeandalucia.es
Historia del artículo:
Recibido el 5 de enero de 2012
Aceptado el 2 de septiembre de 2012
