





3ª PONENCIA
LA PAREJA ESTÉRIL DEL SIGLO XXI
Inducción de la ovulación: hormonas recombinantes
J. Balasch
Departamento de Obstetricia y Ginecología
Facultad de Medicina-Hospital Clínic i Provincial
Casanova, 143
08036 Barcelona
INTRODUCCION
La estimulación del desarrollo folicular ovárico y la inducción de la ovulación constituyen en conjunto la modalidad terapéutica más ampliamente aplicada en el tratamiento de la pareja estéril. Ello se debe al gran auge que han alcanzado las técnicas de reproducción asistida a las que acaban siendo candidatas más del 50% de parejas con problemas reproductivos, ya que tras el fracaso de los diferentes tratamientos estándar para las diferentes causas de esterilidad tanto femeninas como masculinas, la alternativa terapéutica final acaba siendo en la mayoria de casos la inseminación artificial con semen capacitado (IAC) o la fecundación in vitro (FIV) asociada o no a la micromanipulación de los gametos. Ello implica que un gran número de mujeres, ya sea por disfunción ovulatoria o aún siendo normoovuladoras por ser candidatas a técnicas de reproducción asistida, van a ser sometidas a tratamientos inductores de la ovulación o a estimulación (poli)folicular ovárica.
La introducción en estos años recientes de nuevos fármacos encaminados a promover el desarrollo folicular ha constituido uno de los mayores avances en el campo de la endocrinología reproductiva permitiendo así que la inducción y estimulación de la ovulación se haya convertido en una de las áreas más gratificantes para el especialista en reproducción. En este sentido son de destacar las ventajas que han supuesto por ejemplo la introducción de los agonistas de la GnRH en los programas de FIV y lo que puede representar en un futuro inmediato el empleo de los antagonistas; el empleo de las nuevas pautas de inducción de la ovulación en esquemas de pauta lenta; y la disponibilidad de preparados gonadotróficos monohormonales. En relación a este último aspecto hay de destacar la reciente introducción en la farmacopea de las denominadas gonadotrofinas recombinantes que en un futuro no muy lejano han de constituir la única forma de preparados comerciales para empleo en clínica(1).
Las gonadotrofinas recombinantes se obtienen por técnicas de ingeniería genética mediante la integración estable del gen que codifica para la hormona en cuestión en el genoma de la célula huésped, habitualmente de mamífero(2). De esta manera el producto obtenido presenta un grado de pureza absoluto y una bioactividad altamente específica, y la disponibilidad de producto es potencialmente ilimitada con una homogeneidad lote a lote de la que no disponen los preparados actuales de origen urinario (Fig. 1)(3). Estas potenciales ventajas de las gonadotrofinas recombinantes atribuibles a su peculiar forma de obtención han permitido mejorar nuestros conocimientos sobre el control gonadotrófico del ciclo genital y evitar los efectos secundarios asociados esencialmente a la existencia de proteínas contaminantes en los productos de origen urinario.

Figura 1.Potenciales ventajas de la aplicación de las gonadotrofinas recombinantes.
En esta artículo se analizan las aplicaciones clínicas de la hormonas recombinantes folículoestimulante (FSH) (Gonal-FR, Ares-Serono; PuregonR, Organon Internacional), luteinizante (LH) (LHadiR, Ares-Serono) y gonadotrofina coriónica humana (HCG) (OvidrelR, Ares-Serono) en el contexto de las tres principales indicaciones para la administración de gonadotrofinas: la inducción de la ovulación en las pacientes del grupo II de la OMS, el tratamiento de la anovulación en mujeres con hipogonadismo-hipogonadotropo (grupo I de la OMS), y la inducción del desarrollo folicular múltiple para reproducción asistida. Las potenciales aplicaciones de las gonadotrofinas recombinantes en el tratamiento de varones subfértiles con hipogonadismo hipogonadotropo(4,5), escapan a los objetivos del presente artículo.
CARACTERISTICAS BASICAS DE LAS HORMONAS GLICOPROTEICAS
Las tres gonadotrofinas FSH, LH y HCG pertenecen a la familia de las hormonas glicoproteicas de las cuales forma parte también la hormona estimulante del tiroides (TSH). Estas cuatro hormonas son moléculas heterodiméricas constituidas por por dos subunidades (cadena a y cadena ß) unidades por enlaces no covalentes(6). Cada una de las dos subunidades por si misma carece de actividad biológica y por tanto es esencial la formación del heterodímero para que exista actividad endocrina. La cadena a es específica de especie y tiene por tanto una secuencia de aminoácidos idéntica para todas las hormonas de la familia. Por el contrario, la subunidad ß es estructuralmente única para cada hormona y determina la especificidad biológica de la molécula(7).
Con todo, las subunidades ß de las diferentes especies comparten una homología parcial en la secuencia de aminoácidos lo que sugiere que muy probablemente proceden de un precursor común(8). Ello explica que la LH y la HCG de una misma especie puedan unirse a, y activar, los mismos receptores gonadales ya que sus cadenas ß son muy similares. Sin embargo, existen variaciones en el grado de glicosilación de las moléculas que crea un espectro de diferencias en cuanto a su carga electroforética, peso molecular, bioactividad y vida media(9). En efecto, cada hormona glicoproteica contiene varias cadenas colaterales hidrocarbonadas que difieren de forma importante y tienen un carácter/actividad ácido o básico. Estas cadenas hidrocarbonadas incluyen manosa, galactosa, fructosa, glucosamina, N-acetil galactosamina, y ácido siálico. Es este último el elemento más importante y el que tiene un mayor impacto sobre el metabolismo y la actividad biológica de estas hormonas(10). Así, se acepta que las glicoformas de gonadotrofinas con mayor carga de ácido siálico circulan durante un período de tiempo más prolongado en comparación con las formas más básicas y la vida media in vivo es un factor que puede contribuir a determinar la biopotencia global de la hormona. Sin embargo, las isoformas más acídicas tienen también un menor grado de afinidad por el receptor y una menor bioactividad in vitro que las formas menos acídicas(11).
El tipo y distribución de las isoformas (glicoformas) hormonales parecen estar bajo control endocrino ya que varían con la edad de la mujer, el estado menopáusico y la fase del ciclo menstrual probablemente dependiendo de la frecuencia de pulsos de secreción de la GnRH y del perfil esteroideo existente en cada momento(12-15). Así, las mujeres jóvenes tienden a tener isoformas de FSH y LH más básicas (con menos cadenas de ácido siálico) que las mujeres de edad reproductiva avanzada. A su vez, estudios cromatográficos indican que en fase folicular inicial y media existen isoformas más ácidas que a mitad de ciclo, mientras que las formas aún más ácidas se detectan en fase lútea. Esto se debe a una selección selectiva de isoformas secretadas por parte de la hipófisis y el significado biológico de este patrón de secreción de isoformas se desconoce. Se interpreta como un efecto relacionado en parte con los niveles de esteroides circulantes y mediado quizá por cambios en el patrón de secreción de GnRH hipotalámica. Los niveles crecientes de estradiol inducen la aparición de isoformas básicas mientras que en fase lútea los niveles crecientes de progesterona se asocian a una mayor proporción de formas más ácidas. Tras la menopausia aparecen formas más ácidas de FSH y LH tanto en la hipófisis como en la circulación. Estas isoformas acídicas se encuentran también en mujeres con menopausia prematura y en mujeres con síndrome de Turner. Dichas isoformas tienen una vida media más prolongada en la circulación debido a su mayor carga negativa lo que contribuye a la elevación de los niveles de gonadotrofinas circulantes propios del estado climatérico.
Finalmente hay que señalar que también para la HCG en el embarazo se detectan cambios en el perfil de isoformas secretadas de manera que a partir de la semana 13 las moléculas de HCG tienen una menor actividad biológica asociada a un carácter menos ácido, un aparente menor peso molecular y una vida media más corta en circulación que la HCG del primer trimestre del embarazo. Aún cuando el significado biológico de esto se desconoce se postula que esta disminución de actividad biológica en la molécula secretada coincide con la fase en que la placenta se convierte en la principal fuente de secreción esteroidea y la HCG pierde gradualmente su influencia gonadotrófica en la madre(15).
FSH RECOMBINANTE
Existen dos preparados de FSH recombinante que han sido registrados como «folitropina alfa» (Gonal-FR) y «folitropina beta» (PuregonR). La nomenclatura de estos productos recombinantes no tiene nada que ver con las subunidades a y ß de la molécula de FSH y se refiere simplemente al orden cronológico de comercialización. Ambas folitropinas son estructuralmente idénticas a la FSH nativa y cada una de ellas contiene por tanto las dos cadenas a y ß que deben tener un «core» o núcleo proteico idéntico por cuanto se producen tras la transferencia de plásmidos que contienen los dos genes que codifican para las dos subunidades de la FSH en el genoma de líneas celulares de ovario de hembra del hámster chino(2,16). Con todo, se ha señalado que dependiendo de las condiciones de cultivo empleadas y de los enzimas glicosilantes presentes durante la síntesis de la hormona la composición de carbohidratos de la molécula resultante podría variar. Así, el patrón de isoformas de FSH en la folitropina beta es discretamente más ácido (y parecido a la isoforma presente en fase folicular media del ciclo natural) que el de la folitropina alfa (cuyo patrón de isoforma corresponde al de la fase folicular avanzada)(16,17). Si la carga terminal de ácido siálico determina la vida media in vivo y la actividad biológica dependiera únicamente de ello, podría inferirse que esto implica una potencial mayor biopotencia para el preparado con una ligera mayor proporción de isoformas ácidas. Sin embargo esto no es tan sencillo y hay diferentes razones que cuestionan la validez de este razonamiento.
En primer lugar, si bien tal como se ha señalado anteriormente la vida media de la molécula in vivo es un factor que puede contribuir a determinar la biopotencia global de la hormona, esto no lo es todo. En efecto, las formas menos ácidas tienen también una mayor actividad a nivel del receptor y una afinidad por el mismo tres veces superior que las formas ácidas y su bioactividad post-receptor in vitro es también muy superior, por por lo que a pesar de tener una vida media más corta pueden ser realmente más potentes que las forma de más larga duración(11,17). Esto lo demuestra el hecho de que las moléculas de FSH recombinante carentes totalmente de ácido siálico tienen una mayor actividad in vitro que las formas con una elevada carga de este ácido si bien en estos casos extremos aquellas carecen de actividad biológica in vivo debido a la rapidez de su eliminación del torrente sanguíneo(18).
En segundo lugar, tal como se comentó antes, las mujeres en edad reproductiva joven tienen un predominio de formas básicas en comparación con mujeres de más edad. Si esto se correlaciona con la experiencia clínica por la que sabemos que en general las mujeres más jóvenes desarrollan más folículos maduros en un menor período de tiempo y con una menor dosis estimulatoria de gonadotrofinas, podría interpretarse como que las isoformas menos ácidas, aún cuando tendrían una vida media más corta en la circulación, poseerían en cambio una mayor actividad y eficacia biológica para regular la estimulación folicular.
Tercero, de acuerdo con aquel criterio de a mayor carga ácida (menor pH) más biopotencia, la FSH urinaria, especialmente la altamente purificada, debería ser el preparado de FSH con mayor potencia biológica ya es el que presenta un menor pH. Sin embargo esto parece estar en contradicción con la experiencia clínica dado que estudios recientes tanto en mujeres con síndrome de ovarios poliquísticos(19,20) como en pacientes sometidas a FIV bajo supresión hipofisaria con análogos de la GnRH(21,22) demuestran que la FSH recombinante es más eficaz (en base a su mayor biopotencia) que la FSH «pura»(20,21) y que la FSH altamente purificada(19,22).
Por tanto, queda claro de todo lo anterior que no se puede establecer una sencilla correlación entre acidez relativa (carga de ácido siálico), vida media y actividad biológica in vivo de la molécula de gonadotrofina. Como corolario a todo ello hay que destacar los resultados de un estudio muy reciente en el que se compararon por primera vez los resultados obtenidos con folitrofina alfa (Gonal-FR) y con folitrofina beta (PuregonR) en pacientes sometidas a FIV bajo supresión hipofisaria con análogos de la GnRH(23). Los resultados de este estudio comparativo, randomizado, en el que el médico desconocía el producto que se estaba administrando, se resumen en la figura 2. Con una dosis total idéntica de FSH empleada en ambos grupos de tratamiento, el número de ovocitos obtenido fue también el mismo, así como el porcentaje de pacientes que pudieron tener transferencia embrionaria. Dado que el estudio incluía 22 pacientes por grupo, son necesarias nuevas investigaciones con una mayor númeo de casos para determinar si las aparentes diferencias observadas en cuanto a la tasa de embarazos y la tolerancia local entre ambos preparados tienen significado clínico o no.
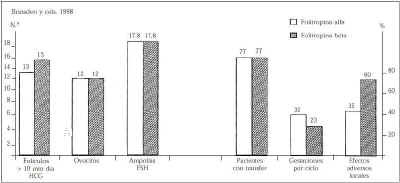
Figura 2.Resultados de un estudio comparativo entre la folitropina alfa y la folitrofina beta para la estimulación folicular múltiple en pacientes sometidas a fecundación in vitro (Brinsden y cols. 1998)(23).
Aplicaciones clínicas de la FSH recombinante
Tanto estudios in vitro o experimentales en animales como la experiencia clínica demuestran claramente que la FSH es el único factor necesario para inducir el crecimiento folicular en diferentes especies animales incluidos los humanos, pero dichas investigaciones demuestran también que la LH es necesaria para que la madurez folicular sea completa y los ovocitos adquieran la capacidad de ser fertilizados normalmente(24-29). Por otra parte, la producción de unas tasas de estradiol normales y la obtención de un endometrio adecuado para la implantación requieren la contribución de la LH tal como lo demuestran los casos de mujeres con déficits aislados de gonadotrofinas tratadas con FSH recombinante(26,29,30,31). Por tanto, las mujeres anovuladoras del grupo I de la OMS necesitarán la administración tanto de FSH como de LH ya que carecen de niveles endógenos suficientes de ésta última hormona.
Por el contrario, existe consenso general de que cuando la LH está ya de por si elevada (como en el caso de las pacientes con síndrome de ovarios poliquísticos) o de forma más general cuando la secreción endógena de LH es normal (caso de las mujeres anovuladoras del grupo II de la OMS), la administración de FSH pura es conceptualmente mejor(28,32). En las pacientes con ovarios poliquísticos la administración de FSH recombinante en un régimen de pauta lenta induce el crecimiento folicular, estimula la secreción de inhibina y de estradiol de manera suficiente para la obtención de una secreción mucosa cervical y de un endometrio adecuados para la consecución del embarazo, promueve la receptividad folicular a la LH/HCG, y se asocia a una disminución significativa de los niveles endógenos de LH durante la fase folicular (evitando así la potencial acción deletérea de una LH elevada sobre la calidad del ovocito resultante)(33).
Cuando se comparan la FSH recombinante y la FSH de origen urinario para la inducción de la ovulación en las pacientes del grupo II de la OMS, no se observan diferencias en las tasas de embarazos observados entre los dos tipos de tratamiento, oscilando dichas tasas entre un 20 y un 33% de gestaciones por ciclo tratado y con un índice acumulativo de embarazos del 40-48% de pacientes tras al menos tres ciclos de tratamiento(5,6,20,34). Es de destacar, por otra parte, que dos estudios recientes demuestran que la FSH recombinante es más eficaz que la FSH urinaria para la inducción de la ovulación en este tipo de pacientes tal como se demuestra por la significativa menor duración del tratamiento, de la dosis total de FSH empleada y de la dosis umbral media de FSH (dosis media diaria efectiva) en el grupo de tratamiento con FSH recombinante versus la FSH urinaria altamente purificada (Fig. 3)(19) o la FSH «pura»(20). El hecho de que esta reducción de la dosis de hormona necesaria no se observara en los primeros estudios al respecto(5,34) se explica porque en dichos ensayos se empleó la hormona recombinante (folitrofina alfa) presentada en forma de viales frente a la FSH urinaria que va contenida en ampollas. Posteriormente se comprobó que la presentación en viales hace que se pierda parte del producto activo al quedar retenido en el vial. Actualmente la FSH recombinante se comercializa en forma de ampollas.

Figura 3.Resultados comparativos (mediana y rango) del número total de ampollas y días de estimulación ovárica con gonadotrofinas empleados con FSH altamente purificada y con FSH recombinante (folitropina alfa) en el mismo grupo de pacientes estériles anovuladoras del grupo II de la OMS tratadas en ciclos consecutivos. Tomado de Balasch y cols. 1998(19).
De lo anterior se deduce por tanto que si bien ambos preparados gonadotróficos son clínicamente útiles, la FSH recombinante puede tener una mayor biopotencia in vivo en comparación con la FSH urinaria. Ello se evidencia especialmente en situaciones tales como el ovario poliquístico en las que existe una disrupción en la acción de la FSH a nivel intraovárico, mientras que en mujeres normoovuladoras sometidas a estimulación del desarrollo folicular para IAC durante un corto período de tiempo a partir del día séptimo del ciclo cuando ya se ha producido la selección del folículo dominante, no se observan diferencias entre la FSH recombinante y la urinaria altamente purificada(19). Además de la reducción de la dosis total y la dosis umbral, otro argumento en favor de esta mayor biopotencia de la FSH recombinante es el estudio de la correlación existente entre el índice basal de LH/FSH y la duración y dosis del tratamiento con FSH. Con la FSH urinaria altamente purificada se observó una correlación directa entre aquellos parámetros, indicando así que a mayor «intensidad» del trastorno endocrinológico mayor cantidad y duración del tratamiento corrector se necesita. Pero cuando se emplea la FSH recombinante en el mismo grupo de mujeres que fue tratada con la FSH urinaria, dicha correlación no existe. Con la FSH recombinante, tanto la mediana como el rango de variación interindividual de la cantidad y duración del tratamiento con FSH fueron significativamente menores que con FSH altamente purificada(19). Podría decirse pues que la FSH recombinante «iguala» a las mujeres con síndrome de ovarios poliquísticos en cuanto a una de sus características endocrinas preeminentes, o dicho de otra manera, es eficaz independientemente del índice LH/FSH basal de la paciente debido a su mayor bioactividad.
No existe una explicación definitiva para la supuesta mayor bioactividad de la FSH recombinante en comparación con la FSH urinaria pero pueden plantearse diferentes hipótesis. En primer lugar, el establecimiento de la biopotencia de las gonadotrofinas se basa en un bioensayo in vivo (el bioensayo de Steelman-Pohley) que ya de por sí es poco sensible y muy impreciso (variación inherente al ensayo del 40%) y tiene una pobre correlación con la respuesta clínica observada en humanos(22,32). Los preparados de gonadotrofinas obtenidos con técnicas recombinantes y que tienen una composición de isoformas constante podrían ser calibrados por medio de bioensayos in vitro recombinantes, eventualmente basados en los receptores gonadotróficos humanos, que son muy fiables, constantes en sus resultados y mucho más precisos que los bioensayos tradicionales(32). Esto comportaría un asignamiento mucho más preciso de la biopotencia y una variabilidad interindividual mucho menor en cuanto a la respuesta terapéutica observada. Por el contrario, diferencias mínimas en el perfil de isoformas de FSH y una considerable variabilidad lote-a-lote en los preparados de origen urinario puede resultar en una diferente regulación de la esteroidogénesis y en variaciones de la FSH bioactiva circulante que estimula la gónada de forma real y efectiva(35,36). Finalmente, otros factores que se postulan para explicar la mayor biopotencia de la FSH recombinante en comparación con la FSH urinaria incluye sutiles diferencias en el perfil de glicoformas o la presencia de proteínas contaminantes en el producto urinario que inhibien la acción de la FSH(21,22).
Además de la mayor biopotencia y de acuerdo con hallazgos recientes, la FSH recombinante podría tener también un mejor perfil de bioactividad in vivo en comparación con la FSH urinaria altamente purificada. En efecto, un estudio in vitro muy reciente ha demostrado que las células de la granulosa del folículo ovárico humano son altamente respondedoras a la FSH recombinante tal como se demuestra por el incremento observado en la producción de esteroides e inhibina(37). Además en dicho estudio se observó una estrecha correlación entre la producción esteroidea y de inhibina indicando así que la regulación de la síntesis y secreción de estas hormonas están íntimamente relacionadas(37).
Hay que recordar que tanto el estradiol como la inhibina A son secretados por las células de la granulosa y ambos son indicadores de crecimiento folicular si bien recientemente se ha indicado que la inhibina A podría reflejar mejor el grado de madurez y funcionalismo foliculares(38-40). Existe una estrecha correlación entre la FSH y la inhibina A durante el ciclo ovárico y la inhibina puede actuar dentro del ovario promoviendo el desarrollo folicular(38,40). Así se ha demostrado que la producción de inhibina por las células de la granulosa estimuladas por la FSH puede verse potenciada por los andrógenos(41). A su vez, los estrógenos pueden ejercer un efecto indirecto sobre la síntesis de inhibina a través del estímulo de la síntesis androgénica en las células tecales(42). Además, la inhibina producida por las células de la granulosa puede actuar como regulador paracrino estimulando la producción androgénica por el compartimiento tecal(43), lo que a su vez proporciona sustrato para la síntesis estrogénica en la granulosa.
En la clínica y en mujeres con síndrome de ovarios poliquísticos, nosotros observamos una correlación directa entre las concentraciones plasmáticas de estradiol y de inhibina A el día de la inyección de la HCG en los ciclos de tratamiento con FSH recombinante pero no en los tratados con FSH altamente purificada en la misma mujer(19). Por tanto, este perfil endocrinológico folicular aparentemente más fisiológico es otro argumento en favor de un mejor comportamiento bioactivo de la FSH recombinante en comparación con la FSH altamente purificada. Y en conjunto, todos los datos anteriores señalan a la FSH recombinante como el tratamiento de elección en las pacientes con anovulación del grupo II de la OMS. En este sentido es de destacar que a pesar de su mayor biopotencia y de ser este tipo de pacientes las más susceptibles de desarrollar un síndrome de hiperestimulación ovárica, la FSH recombinante no implica un factor de riesgo añadido para el desarrollo del síndrome si se emplea según los protocolos de pauta lenta a baja dosis(44).
En consonancia con lo anterior, la FSH recombinante se ha mostrado eficaz para la estimulación polifolicular en pacientes sometidas a FIV sin desensibilización hipofisaria en las que existen por tanto unos niveles normales de LH endógena(45). Sin embargo, en el momento actual la mayoría de programas de FIV emplean la supresión hipofisaria con análogos de la GnRH concomitantemente a la estimulación ovárica con gonadotrofinas y por tanto es en esta situación donde ha de valorarse la eficacia de la FSH recombinante.
En pacientes tratadas mediante FIV bajo supresión hipofisaria con análogos de la GnRH, dos estudios recientes(21,22) demuestran que la FSH recombinante es más eficaz que la FSH urinaria «pura»(21) o altamente purificada(22) ya que con una menor dosis total de gonadotrofina administrada y menos días de tratamiento, la respuesta ovárica en términos de desarrollo folicular, niveles de estradiol y número de ovocitos obtenidos, fue significativamente superior en el grupo de tratamiento con FSH recombinante. Los resultados de la FIV fueron también superiores con este tipo de preparado de FSH cuando se valoran el número y calidad de los embriones obtenidos. Esto a su vez, podría explicar que cuando se analizan conjuntamente varios estudios en los que se compararon ambas preparaciones de FSH, se encuentre una tasa de embarazos significativamente superior para la FSH recombinante (22,9% gestaciones evolutivas por ciclo) que para la FSH urinaria «pura» (17,9% de embarazos evolutivos)(46). Sin embargo, globalmente estas tasas de embarazo parecen algo bajas, en especial para la FSH «pura» si se considera el índice de gestaciones evolutivas obtenido en el estudio en que se comparó la FSH recombinante (34% de embarazos por ciclo) con la FSH altamente purificada (32% de gestaciones por ciclo)(22). De nuevo, el hecho de que no se observaran diferencias en la cantidad de FSH utilizada en estudios previos(47) en que se compararon la FSH recombinante y la FSH «pura» en FIV, se puede explicar porque la FSH recombinante se utilizó en forma de viales en vez de ampollas tal como se comentó antes.
En estos estudios anteriores, sin embargo, la FSH recombinante se comparó con la FSH urinaria con lo que el aporte exógeno de LH es nulo o prácticamente nulo en ambos grupos de tratamiento. En el momento actual el uso de análogos de la GnRH para la supresión hipofisaria en los ciclos de FIV es una práctica generalizada, siendo la forma habitual de uso el «protocolo largo» en el que no se inicia la estimulación ovárica con gonadotrofinas hasta haberse conseguido la hipofisectomía médica. En estas condiciones transitorias de hipogonadismo hipogonadotropo endógeno, la experiencia clínica demuestra que el estímulo con FSH recombinante exógena parece suficiente en la mayoría de mujeres para conseguir una adecuada respuesta ovárica en los ciclos de FIV. La interpretación a este hecho es que en la mayoría de pacientes quedan unos niveles residuales de LH bioactiva suficientes para que se produzca la esteroidogénesis, lo que se explicaría por el hecho fisiológico de que basta la ocupación de < 1% de todos los receptores gonadales de LH para obtener una respuesta esteroidogénica máxima(48,49). Por otra parte se sugiere que los niveles de LH requeridos para apoyar a la FSH en la síntesis esteroidea son bajos, de unas 2 UI/l(50).
Pero frente a lo anterior, hay datos en la literatura que apoyan la posibilidad de un déficit de respuesta folicular ovárica en mujeres con ciclos regulares que son sometidas a tratamiento con preparados monohormonales de FSH altamente específicos bajo hipofesectomía médica. Son los siguientes:
Primero, tanto la GnRH como sus análogos pueden tener acciones extrahipofisarias, incluyendo el ovario, en el que se ha demostrado la existencia de receptores específicos(51-53).
Segundo, el nivel de supresión hipofisaria alcanzado (y por tanto el nivel residual de LH existente post-inhibición) varía dependiendo del tipo de análogo utilizado, de la pauta de administración del mismo y de la especial susceptibilidad de la paciente(50,54-57). En otras palabras, que el grado de supresión hipofisaria que se obtiene con los análogos de la GnRH puede variar de un individuo a otro dependiendo no sólo del preparado utilizado y su potencia respectiva, sino también de la vía y dosis administrada y ello puede influir en los resultados de la FIV. De hecho, existen diferencias en los resultados de la FIV según la potencia del análogo de la GnRH utilizado(58). Pero además, puede haber marcadas diferencias en el grado de supresión hipofisaria obtenido de un individuo a otro aún empleando la misma formulación y dosis de análogo(55,56).
Tercero, cuando se emplean los agonistas de la GnRH en régimen de protocolo largo, se requiere una dosis significativamente mayor de FSH recombinante para la estimulación folicular en FIV que cuando se aplica el protocolo corto, lo que indica que el nivel residual de LH post-inhibición influye sobre la capacidad de respuesta ovárica al estímulo con gonadotrofinas exógenas(50).
Cuarto, las determinaciones de los niveles de LH inmunoactiva en suero tienen un valor muy limitado, si es que tienen alguno, para identificar si una paciente tendrá suficiente secreción endógena de LH para responder adecuadamente a la estimulación con FSH sola(59,60).
Por último, diferentes estudios indican que los niveles máximos de estradiol obtenidos durante la estimulación folicular ovárica bajo supresión hipofisaria con análogos de la GnRH para FIV, son superiores cuando se emplean las HMG frente a preparados monohormonales de FSH(61-63).
Por tanto todo lo anterior significa que hay bases biológicas plausibles para explicar dos hechos observados en la práctica clínica diaria: a) la falta de inhibición hipófiso-ovárica en algunas pacientes frente a una posología estándar de análogo de la GnRH en un programa de FIV; y b) el déficit de desarrollo folicular para un ciclo de FIV en un subgrupo de mujeres catalogadas como «low-responders» (respuesta más lenta o finalmente insuficiente), que tienen unas características endocrinas basales similares a las de las normorespondedoras y que reciben la misma dosis de FSH. La existencia de este último subgrupo de mujeres podría explicarse por los efectos directos sobre el ovario de los análogos de la GnRH a los que hemos hecho mención, pero implica también la posibilidad de que la supresión demasiado profunda de LH haga insuficiente el tratamiento exógeno con FSH sola tal como sugieren algunos estudios experimentales(64,65).
En la clínica práctica y apoyando esta posibilidad es de destacar un estudio muy reciente que demuestra que las pacientes de FIV en las que se alcanza una profunda depresión de los niveles plasmáticos de LH (< 0,5 UI/l) a mitad de la fase de estimulación folicular con FSH pura y buserelin, tienen un desarrollo polifolicular normal pero las cifras de estradiol absolutas y por cada folículo desarrollado, el número de ovocitos obtenidos, la tasa de fertilización y el número de embriones conseguidos son inferiores a los obtenidos en las pacientes con niveles mayores de LH(66).
Así pues, tal como indica Hillier(28), «está por establecer si la suplementación con LH recombinante en las pautas de tratamiento con FSH recombinante bajo análogos supondrá o no alguna ventaja. El potencial beneficio de añadir la LH podría depender del grado en que los niveles plasmáticos endógenos de LH quedan suprimidos por el agonista». Hoy por hoy, hay que aceptar que la desensibilización hipofisaria con análogos asociada a la estimulación folicular con FSH recombinante es una pauta aplicable a la mayoría de mujeres con ciclos regulares sometidas a FIV, si bien es posible que en algunas pacientes la respuesta ovárica esté algo por debajo de lo normal cuando se aplica este pauta. No existen datos que permitan identificar a priori a este subgrupo de pacientes que posiblemente representan tan sólo alrededor del 5% de casos y que sólo se manifiestan por tener una respuesta folicular y unos niveles de estradiol por debajo de la media de las mujeres. Por ello, pueden pasar desapercibidas en el cómputo general de los resultados de un programa de FIV. Por otra parte hasta ahora se carecía de la posibilidad de tratarlas suplementariamente con un preparado específico de LH y habrá que esperar por tanto a futuros estudios para aclarar el potencial beneficio de la adición rutinaria de LH recombinante a la FSH recombinante.
LH RECOMBINANTE
La LH recombinante (LHadiR, Ares-Serono) ha estado disponible para la práctica de ensayos clínicos desde 1993. Los primeros estudios con esta hormona se realizaron en primates no humanos y consistieron en una investigación farmacocinética encaminada a valorar y comparar la LH hipofisaria, urinaria y recombinante humanas(67). Tras la administración i.v. de cada una de las tres formas de la hormona se constató que las concentraciones circulantes alcanzadas en relación al tiempo desde la inyección del producto, el índice de aclaramiento, el volumen de distribución y la vida media del principio activo fueron similares para las tres preparaciones de LH. Por otra parte, se demostró que la LH recombinante era una alternativa válida a la LH nativa y lógicamente a la HCG en su capacidad para desencadenar el fenómeno ovulatorio en el folículo de los primates(68).
Por otro lado, estudios en primates tratados con antagonistas de la GnRH demostraron que la adición de LH recombinante a la FSH recombinante no sólo normaliza la capacidad de secreción esteroidea ovárica sino que además acorta significativamente los días de tratamiento con FSH recombinante necesarios para lograr el desarrollo y maduración foliculares(69). A todo esto le siguió la demostración, por primera vez, de la eficacia y seguridad de la LH recombinante cuando se emplea en humanos para inducir la ovulación y conseguir el embarazo en mujeres con hipogonadismo hipogonadotropo(70-72).
La consecuencia lógica de todo lo anterior era establecer cuales son las necesidades mínimas (dosis mínima efectiva) de LH en mujeres con anovulación del grupo I de la OMS (hipogonadismo hipogonadotropo), la aplicación clínica princeps de la LH recombinante. Esto se ha llevado a cabo en un estudio publicado muy recientemente(73) en el que 38 pacientes estériles anovuladoras con hipogonadismo hipogonadotropo fueron tratadas con una dosis fija de 150 UI de FSH recombinante (Gonal-FR) y distribuidas de forma randomizada en cuatro grupos para recibir LH recombinante (LHadiR) a dosis diarias de 0 UI, 25 UI, 75 UI, ó 225 UI. Las principales conclusiones de dicho estudio, que establece claramente la eficacia clínica de la LH recombinante asociada a la FSH recombinante para inducir un crecimiento y función folicular normales en aquel tipo de pacientes, fueron las siguientes:
1. La LH recombinante promueve en forma dosis-dependiente la secreción ovárica de androstendiona y de estradiol por los folículos en crecimiento estimulados por la FSH recombinante. Esto es, las concentraciones séricas de estradiol el último día de tratamiento con FSH fueron de 18 ± 1 pg/ml, 53 ± 25 pg/ml, 379 ± 159 pg/ml, y 665 ± 331 pg/ml, mientras que para la androstendiona fueron de 103 ± 25 ng/dl, 146 ± 37 ng/dl, 183 ± 37 ng/dl, y 192 ± 37 ng/dl, para las pacientes tratadas con 0, 25, 75, y 225 UI diarias de LH recombinante respectivamente.
2. La LH recombinante aumenta la sensibilidad ovárica a la FSH tal como se demuestra por la proporción de pacientes que presentaron desarrollo folicular tras la administración de la dosis fija de FSH y que fueron 12,5%, 43%, 78%, y 80% de las pacientes tratadas con 0, 25, 75, y 225 UI diarias de LH respectivamente.
3. La LH recombinante aumentó la capacidad de luteinización de los folículos tras la administración de HCG.
4. La dosis diaria de 75 UI de LH recombinante fue eficaz en la mayoría de mujeres (80% de ciclos) para promover un desarrollo folicular óptimo definido por la existencia de 31 folículo de 317 mm de diámetro con un estradiol 3110 pg/ml, una progesterona plasmática en fase lútea media 378 ng/ml, y un crecimiento endometrial apropiado.
Algunas pacientes pueden requerir hasta 225 UI de LH recombinante diarias para que se cumplan las anteriores premisas, pero es interesante destacar que en el grupo global de pacientes que recibieron la dosis diaria de 225 UI hubo una tendencia al desarrollo de un menor número de folículos lo que podría estar de acuerdo con la teoría del «techo de la LH»(28) según la cual la LH es necesaria para el desarrollo del folículo pero por encima de ciertos niveles promueve la atresia folícular. Este es un hecho que parece bien constatado en el ciclo natural pero que requeriría confirmación para la terapéutica con gonadotrofinas exógenas, especialmente teniendo en cuenta el punto 5 siguiente.
5. Aunque la dosis diaria de 75 UI de LH recombinante fue suficiente (dosis mínima eficaz) para promover el desarrollo folicular en la mayoría de pacientes del grupo I de la OMS, los niveles séricos de LH permanecieron por debajo de los límites de cuantificación del ensayo (1 UI/l). Sólo en tres de las diez pacientes tratadas con 225 UI se detectaron valores de LH en suero de 1.3, 1.4, y 1.6 UI/L respectivamente. Esto indica pues las importantes limitaciones de las determinaciones hormonales de LH basadas tan sólo en la inmunoreactividad y no en la bioactividad.
6. Los resultados clínicos obtenidos en este reciente estudio(73) empleando la asociación de FSH y LH recombinantes en pacientes con hipogonadismo hipogonadotropo son similares a los obtenidos con HMG o con infusión pulsátil de GnRH en términos de tasas de ovulación y de embarazo por ciclo de tratamiento en este tipo de pacientes (Tabla 1).
Tabla 1 Resultados de diferentes tipos de tratamiento para la inducción de la ovulación en pacientes con hipogonadismo hipogonadótropo | |||
| Autor, año | Fármaco: dosis inicial y (vía de administración) | Ovulaciones/ciclo (%) | Embarazos/ciclo (%) |
| Couzinet, 1988 | HMG: 225 Ul/d (i.m.) | 100 | ? |
| Shoham, 1991 | HMG: 75 Ul/d (i.m.) | 89 | ? |
| Martin, 1993 | HMG: 150 Ul/d (i.m.) | 97 | 25 |
| Balen, 1994 | HMG: 75 Ul/d (i.m.) | 84 | 18 |
| Martin, 1993 | GnRH: 75-250 ng/kg (bolus i.v.) | 93 | 29 |
| Skarin, 1993 | GnRH: 1-40 mcg/bolus (i.v., s.c.) | 83 | 27 |
| Filicori, 1994 | GnRH: 1,25-20 mcg/bolus (i.v.) | 79 | 22 |
| Braat, 1991 | GnRH: 5-20 mcg/bolus (s.c.) | 68 | 13 |
| Este estudio*, 1988 | FSHr + LHr: 150 + 75 Ul/d (s.c.) | 75 | 29 |
| FSHr + LHr: 150 + 225 Ul/d (s.c.) | 86 | 20 | |
| *The European Recombinant Human LH Study Group. | |||
En las pacientes sometidas a FIV bajo supresión hipofisaria con análogos de la GnRH, la LH recombinante podría tener aplicación para apoyar el crecimiento y desarrollo foliculares asociada a la FSH recombinante si se confirmara, tal como se discutió antes, la existencia de un pequeño subgrupo de pacientes en las que el grado de desensibilización hipofisaria es tan intenso que los niveles de LH endógena bioactiva residuales son insuficientes para el logro de un desarrollo folicular múltiple y una esterodogénesis adecuados(66). Pero además de este mecanismo indirecto de déficit de LH por acción de los análogos, existen también evidencias experimentales en la literatura que apoyan el posible papel beneficioso directo del aporte exógeno de LH no sólo sobre la calidad de la foliculogénesis, sino también sobre la del ovocito y del embrión(74-77). Si ello se confirmara en los humanos, estaría justificado el empleo rutinario de LH recombinante asociada a la FSH recombinante en FIV.
Pero cabe además una tercera potencial aplicación clínica para la LH recombinante en pacientes sometidas a FIV y que podría constituir una indicación no selectiva sino generalizada. Estudios de nuestro grupo han demostrado que la patogenia del síndrome de hiperestimulación ovárica grave se asocia de forma constante a un estado de vasodilatación arterial periférica que constituye un hecho fundamental en el desarrollo de las manifestaciones clínicas del cuadro(78-80). Estas pacientes presentan un estado de circulación hiperdinámica caracterizado por hipotensión arterial, aumento del gasto cardíaco y disminución de las resistencias vasculares periféricas, indicando así la existencia de la vasodilatación arteriolar periférica. Ello conduce a una hiperactividad de los sistemas vasoactivos endógenos como parte de una respuesta homeostática generalizada encaminada a mantener la función circulatoria y la presión arterial. Sin embargo, el aumento de actividad simpática y el incremento de los niveles de aldosterona y hormona antidiurética promueven la retención de sodio y agua que contribuye a la formación de edema y ascitis.
Pero aún más interesante es el hecho siguiente. En otro estudio más reciente(81) hemos demostrado que esta disfunción circulatoria que caracteriza al síndrome de hiperestimulación ovárica grave es un fenómeno universal (aunque lógicamente de menor intensidad) en las pacientes sometidas a hiperestimulaticón ovárica controlada para FIV sugiriendo así que la forma grave del síndrome clínico sería la expresión extrema de dicho trastorno. Es importante destacar además que si bien el incremento de los niveles de estradiol durante los ciclos de FIV se asocia a modificaciones hemodinámicas significativas (aumento del gasto cardíaco y disminución de las resistencias vasculares periféricas), la disfunción circulatoria que caracteriza al síndrome de hiperestimulación ovárica grave con las alteraciones neurohormonales que comporta, es claramente independiente del inicio de la hiperestrogenemia. Por el contrario, parece claro el papel patogénico que juega la inyección de la dosis ovulatoria de HCG en la disfunción circulatoria observada de forma constante en las pacientes de FIV. Esto se basa en los hechos siguientes:
1. Todas las pacientes incluidas en nuestro estudio(81) estuvieron asintomáticas a lo largo del ciclo de FIV pero desarrollaron la disfunción circulatoria que caracteriza las formas graves del síndrome (disminución de tensión arterial y de resistencias vasculares periféricas, aumento del gasto cardíaco y marcada elevación de los niveles de actitividad renina y concentración de norepinefrina en plasma) siete días tras la administración de la HCG.
2. La disfunción circulatoria tras la inyección de HCG apareció precisamente en el momento en que suelen desarrollarse las manifestaciones clínicas del síndrome de hiperestimulación ovárica severo(82-84).
En relación a lo anterior hay que destacar que la HCG tiene una vida media más larga que la LH, posee un efecto más prolongado sobre los folículos en diferentes estadios madurativos, sus acciones biológicas pueden no ser idénticas a las de la LH, y se asocia a un estímulo suprafisiológico cuando se compara con la LH recombinante para la inducción de la ovulación en el animal de experimentación(85).
Teniendo en cuenta todas estas consideraciones es posible que la disfunción circulatoria observada de forma constante en las pacientes de FIV, y que en su forma extrema puede manifestarse en forma del síndrome de hiperestimulación ovárica grave potencialmente letal para la mujer, pueda evitarse si en vez de HCG se administra LH recombinante para inducir la maduración de los ovocitos. De ser así, la indicación generalizada para el uso de LH recombinante quedaría plenamente justificada.
HCG RECOMBINANTE
En los estudios clínicos de fase I la HCG recombinante (OvidrelR, Ares-Serono) fue administrada a varones y mujeres voluntarios tanto por vía e.v., s.c. ó i.m., comprobándose que sus características farmacocinéticas son superponibles a las de la HCG de origen urinario y sin que existieran prácticamente diferencias entre las tres vías de administración(5).
En el momento actual se está ensayando la administración de HCG recombinante tanto en pacientes sometidas a técnicas de fecundación asistida como a inducción de la ovulación por esterilidad anovulatoria del grupo II de la OMS habiéndose registrado gestaciones en ambos grupos de tratamiento.
Una potencial aplicación clínica específica de la HCG recombinante sería en las pacientes que presentan el denominado «síndrome del folículo vacío» una entidad que implica un coste tanto económico como psicológico y una pérdida de tiempo y esfuerzos tanto para los pacientes como para el equipo médico. Aún cuando su etiopatogenia no está clara en todos los casos, se acepta que en la mayoría de ocasiones es debido a una anomalía de la actividad biológica in vivo de algunos lotes comerciales de la actualmente disponible HCG de origen urinario(86). Por un defecto en el proceso de producción a partir de la orina, la HCG de estos lotes anómalos tendría un índice de aclaramiento plasmático más rápido de lo normal secundariamente a un contenido insuficiente en ácido siálico lo que le impediría ejercer las acciones biológicas esperados(86).
El elevado grado de pureza, la bioactividad altamente específica y la homogeneidad lote a lote que implica el proceso de obtención de las hormonas recombinantes mediante las técnicas de ingeniería genética, podrían evitar ese inoportuno síndrome empleando la HCG recombinante en vez de la urinaria.
BIBLIOGRAFIA
1. Lunenfeld B, Lunenfeld E. Gonadotropic preparations-lessons learned. Fertil Steril 1997;67:812-4.
2. Howles CM. Genetic engineering of human FSH (Gonal-F). Hum Reprod Update 1996;2:172-91.
3. Balasch J, González-Merlo J. Las nuevas gonadotrofinas recombinantes: un avance terapéutico. Prog Obstet Ginecol 1996;39:245-7.
4. Quinton R, Matfin G, Bouloux P, Loumaye E. Successful induction of fertility in a hypogonadotropic male. Lancet 1994;343:733.
5. Loumaye E, Martineau I, Piazzi A y cols. Clinical assesment of human gonadotrophins produced by recombinant DNA technology. Hum Reprod 1996;11(supl 1):95-107.
6. Pierce JG, Parsons TF. Glycoprotein hormones: structure and function. Annu Rev Biochem 1981;50:455-95.
7. Baenziger JU, Green ED. Pituitary glycoprotein hormone oligosaccharides: structure, synthesis, and function of asparagine-linked oligosaccharides on lutropin, follitropin and thyrotropin. Biochim Piophys Acta 1988;947:287-306.
8. Gharib OD, Wierman ME, Shupnik MA, Chin WW. Molecular biology of the pituitary gonadotropins. Endocr Rev 1990;177-98.
9. Ulloa-Aguirre A, Espinoza R, Damian-Matsumura P, Chapel SC. Immunological and biological potencies of different molecular species of gonadotropins. Hum Reprod 1988;3:459-501.
10. Dufau ML, Catt KJ, Tsurahara T. Retention of in vitro biological activities by desialylated human luteinizing hormone and chorionic gonadotropin. Biochem Biphys Res Commun 1971;44:1022-9.
11. Chappel S. The choice of gonadotrophin isoforms in present and future preparations What can we expect? In Gonadotrophin Isoforms, Facts and Future, edited by JA Khan, Ciconia Foundation, Copenhagen, 1997:61-9.
12. Chappel SC. Heterogeneity of follicle stimulating hormone: control and physiological function. Hum Reprod Update 1995;1:479-87.
13. Ulloa-Aguirre A, Midgley AR, Beitins IZ, Padmanabhan V. Follicle stimulating isohormones: Characterization and physiological relevance. Endocr Rev 1995;16:765-87.
14. Burgon PG, Stanton PG, Robertson DM. In vitro bioactivities and clearance patterns of highly purified human luteinizing hormone isoforms. Endocrinology 1996;137:4827-36.
15. Wide L. Isoforms of human gonadotrophins under different physiological conditions. In Gonadotrophin Isoforms, Facts and Future, edited by JA Khan, Ciconia Foundation, Copenhagen, 1997:43-52.
16. Goa KL, Wagstaff AJ. Follitropin alpha in infertility. BioDrugs 1998;9:235-60.
17. Robertson WR. Gonadotrophin isoform pattern in different pharmaceutical preparation. In Gonadotrophin Isoforms, Facts and Future, edited by JA Khan, Ciconia Foundation, Copenhagen, 1997:53-60.
18. Galway AB, Hsueh AJW, Keene JL, Yamoto M, Fauser BCJM, Moime I. In vitro and in vivo bioactivity of recombinant human follicle-stimulating hormone and partially deglycosylated variants secreted by transfected eukaryotic cell lines. Endocrinology 1990;127:93-100.
19. Balasch J, Fábregues F, Peñarrubia J, Creus M, Vidal R, Casamitjana R, Manau D, Vanrell JA. Follicular development and hormonal levels following highly purified or recombinant follicle stimulating hormone administration in ovulatory women and WHO group II anovulatory infertile patients. J Assist Reprod Genet 1998;15:552-9.
20. Coelingh Bennink HJT, Fauser BCJM, Out HJ. Recombinant follicle-stimulating hormone (FSH; Puregon) is more efficient than urinary FSH (Metrodin) in women with clomiphene citrate-resistant, normogonadotropic, chronic anovulation: a prospective, multicenter, assessor-blind, randomized, clinical trial. Fertil Steril 1998;69:19-25.
21. Out HJ, Mannaerts BMJL, Driessen SGAJ, Coelingh Bennink HJT. A prospective, randomized, assessor-blind, multicentre study comparing recombinant and urinary follicle stimulating hormone (Puregon versus Metrodin) in in vitro fertilization. Hum Reprod 1995;10:2534-40.
22. Bergh C, Howles CM, Borg K, Hamberger L, Josefsson B, Nilsson L, Wikland M. Recombinant human follicle stimulating hormone (r-hFSH; Gonal F) versus highly purified urinary FSH (Metrodin HP): results of a randomized comparative study in women undergoing assisted reproductive techniques. Hum Reprod 1997;12:2133-9.
23. Brinsden P, Akagbosu F, Gibbons L, Lancaster S, Gourdon D, Macnamee M, Loumaye E. Gonal-FR versus PuregonR: results of a randomized, assessor-blind, comparative study in women undergoing assisted reproductive technologies. Hum Reprod 13 (Abstract Book 1) (14th Annual Meeting of the ESHRE):1998:70.
24. Couzinet B, Lestrat N, Brailly S, Forest M, Schaison G. Stimulation of ovarian follicular maturation with pure follicle-stimulating hormone in women with gonadotropin deficiency. J Clin Endocrinol Metab 1998;66:552-6.
25. Shoham Z, Balen A, Patel A, Jacobs HS. Results of ovulation induction using human menopausal gonadotropin or purified follicle-stimulating hormone in hypogonadotropic hypogonadism patients. Fertil Steril 1991;56:1048-53.
26. Schoot DC, Coelingh Bennink HJT, Mannaerts BM y cols. Human recombinant follicle-stimulating hormone induces growth of preovulatory follicles without concomitant increase inandrogen and estrogen biosynthesis in a woman with isolated gonadotropin deficiency. J Clin Endocrinol Metab 1992;74:1471-3.
27. Wang XN, Greenwald GS. Human chorionic gonadotropin or human recombinant follicle-stimulating hormone (FSH)-induced ovulation and subsequent fertilization and early embryo development in hypophysectomized FSH-primed mice. Endocrinology 1993;132:1009-2016.
28. Hillier SG. Current concepts of the roles of follicle stimulating hormone and luteinizing hormone in folliculogenesis. Hum Reprod 1994;9:188-91.
29. Balasch J, Miró F, Burzaco I, Casamitjana R, Civico S, Ballescá JL, Puerto B, Vanrell JA. The role of luteinizing hormone in human follicle development and oocyte fertility: evidence from in-vitro fertilization in a woman with long-standing hypogonadotrophic hypogonadism and using recombinant human follicle stimulating hormone. Hum Reprod 1995;10:1678-83.
30. Mannaerts B, Shoham Z, Schoot D y cols. Single-dose pharmacokinetics and pharmacodynamics of recombinant human follicle-stimulating hormone (Org 32489) in gonadotropin-deficient volunteers. Fertil Steril 1993;59:108-14.
31. Shoham Z, Mannaerts B, Insler V, Coelingh Bennink H. Induction of follicular growth using recombinant human follicle-stimulating hormone in two volunteer women with hypogonadotropic hypogonadism. Fertil Steril 1993;59:738-42.
32. Simoni M, Nieschlag E. FSH in therapy: physiological basis, new preparations and clinical use. Reprod Med Rev 1995;4:163-77.
33. Hornnes P, Giroud D, Howles C, Loumaye E. Recombinant human follicle-stimulating hormone treatment leads to normal follicular growth, estradiol secretion, and pregnancy in a World Health Organization group II anovulatory woman. Fertil Steril 1993;60:724-6.
34. Homburg R, Balasch J, Birkhauser M y cols. Efficacy of recombinant human follicle stimulating hormone Gonal-FR for inducing ovulation in WHO group II anovulatory patients Preliminary results of a comparative, multicentre study. In Perspectives on Assisted Reproduction, edited by T Mori et al Ares-Serono Symposia Publications, Roma; 1994:463-7.
35. Mason HD, Mannaerts B, De Leeuw R, Willis DS, Franks S. Effects of recombinant human follicle stimulating hormone on cultured human granulosa cells: comparison with urinary gonadotrophins and actions in preovulatory follicles. Hum Reprod 1993;8:1823-7.
36. Fauser BCJM, Van Heusden AMV. Manipulation of human ovarian function: physiological concepts and clinical consequences. Endocrine Rev 1997;18:71-106.
37. Bergh C, Selleskog U, Hillensjö T. Recombinant human gonadotropins stimulate steroid and inhibin production in human granulosa cells. Eur J Endocrinol 1997a;136:617-23.
38. Groome NP, Illingworth PJ, O''Brien M, Pai R, Rodger FE, Mather JP, McNeilly AS. Measurement of dimerci inhibin B throughout the human menstrual cycle. J Clin Endocrinol Metab 1996;81:1401-5.
39. Klein NA, Illingworth PJ, Groome NP, McNeilly AS, Battaglia DE, Soules MR. Decreased inhibin B secretion is associated with the monotropic FSH rise in older, ovulatory women: A study of serum and follicular fluid levels of dimeric inhibin A and B in spontaneous menstrual cycles. J Clin Endocrinol Metab 1996; 81:2742-5.
40. Lockwood GM, Muttukrishna S, Groome NP, Knight PG, Ledger WL. Circulating inhibins and activin A during GnRH-analogue down-regulation and ovarian hyperstimulation with recombinant FSH for in vitro fertilization-embryo transfer. Clin Endocrinol 1996;45:741-8.
41. Hillier SG, Wickings EJ, Illingworth PJ, Yong EL, Reichert LE Jr, Baird DT, McNeilly AS. Control of immunoactive inhibin production by human granulosa cells. Clin Endocrinol 1991; 35:71-5.
42. Gilling-Smith C, Willis DS, Franks S. Oestradiol feedback stimulation of androgen biosynthesis by human theca cells. Hum Reprod 1997;12:1621-8.
43. Hillier SG, Yong EL, Illingworth PJ; Baird DT, Schwall RH, Mason AJ. Effect of recombinant inhibin on androgen synthesis in cultured human thecal cells. Mol Cell Endocrinol 1991a;75:R1-6.
44. Aboulghar MA, Mansour RT, Serour GI, Amin YM, Sattar MA, ElAttar E. Recombinant follicle-stimulating hormone in the treatment of patients with history of severe ovarian hyperstimulation syndrome. Fertil Steril 1996;66:757-60.
45. Strowitzki T, Kentenich H, Kiessel L, Neulen J, Bilger W. Ovarian stimulation in women undergoing in vitro fertilization and embryo transfer using recombinant human follicle stimulating hormone (Gonal-F) in non-down-regulated cycles. Hum Reprod 1995;10:3097-101.
46. Out HJ, Mannaerts BMJL, Driessen SGAJ, Coelingh Bennink HJT. Recombinant follicle-stimulating hormone (follitropin beta, Puregon*) yields higher pregnancy rates in in vitro fertilization than urinary gonadotropins. Fertil Steril 1997;68:138-42.
47. Recombinant Human FSH Study Group. Clinical assessment of recombinant human follicle-stimulating hormone in stimulating ovarian follicular development before in vitro fertilization. Fertil Steril 1995;63:77-86.
48. Catt KJ, Dufau ML. Spare receptors in rat testes. Nature 1977;244:219-21.
49. Doerr R. Relationship between saturation of LH receptors and steroidogenesis response in isolated rat granulosa cells. Horm Metab Res 1979;11:181-4.
50. Devroey P, Mannaerts B, Smitz J, Coelingh-Bennink H, Van Steirteghem A. Clinical outcome of a pilot efficacy study on recombinant human follicle-stimulating hormone (Org 32489) combined with various gonadotrophin-releasing hormone agonist regimens. Hum Reprod 1994;9:1064-9.
51. Hsueh AJW, Erickson GF. Extrapituitary action of gonadotropin-releasing hormone: direct inhibition of ovarian steroidogenesis. Science 1979;204:854-5.
52. Casper RF, Erickson GF, Rebar RW, Yen SSC. The effect of luteinizing hormone-releasing factor and its agonists on cultured human granulosa cells. Fertil Steril 1982;37:406-9.
53. Bramley TA, Menzies GS, Baird DT. Specific binding of gonadotropin-releasing hormone and an agonist to human corpus luteum homogenates: characterization, properties, and luteal phase levels. J Clin Endocrinol Metab 1985;61:834-41.
54. Vauthier D, Lefebvre G. The use of gonadotropin-releasing hormone analogs for in vitro fertilization: comparison between the standard form and long-acting formulation of D-Trp-6-luteinizing hormone-releasing hormone. Fertil Steril 1989;51:100-4.
55. Maheux R. The use of LHRH analogues in the treatment of leiomyoma uteri. In LHRH Analogues in Gynaecology, edited by Brosens I y cols. Parthenon Publishing, Casterton Hall, 1990; p .127-39.
56. Conn PM, Crowley WF. Gonadotropin-relasing hormone analogues. N Engl J Med 1991;324:93-103.
57. Filicori M, Flamigni C, Cognini G y cols. Comparison of the suppressive capacity of different depot gonadotropin-releasing hormone analogs in women. J Clin Endocrinol Metab 1993;77:130-3.
58. Balasch J, Jové IC, Moreno V y cols. The comparison of two gonadotropin-releasing hormone agonists in an in vitro fertilization. Fertil Steril 1992;58:991-4.
59. Chappel SC, Howles C. Reevaluation of the roles of luteinizing hormone and follicle-stimulating hormone in the ovulatory process. Hum Reprod 1991;6:1206-12.
60. Loumaye E, Engrand P, Howles CM, O''Dea L. Assessment of the role of serum luteinizing hormone and estradiol response to follicle-stimulating hormone on in vitro fertilization treatment outcome. Fertil Steril 1997;67:889-99.
61. Balasch J, Fábregues F, Creus M, Moreno V, Puerto B, Peñarrubia J, Carmona F, Vanrell JA. Pure and highly purified follicle-stimulating hormone alone or in combination with human menopausal gonadotrophin for ovarian stimulation after pituitary suppression in in vitro fertilization. Hum Reprod 1996;11:2400-4.
62. Fried G, Harlin J, Csemickzy G, Wramsby H. Controlled ovarian stimulation using highly purified FSH results in a lower serum oestradiol profile in the follicular phase as compared with HMG. Hum Reprod 1996;11:474-7.
63. Westergaard LG, Erb K, Laursen S y cols. The effect of human menopausal gonadotrophin and highly purified, urine-derived follicle stimulating hormone on the outcome of in vitro fertilization in down-regulated normogonadotrophic women. Hum Reprod 1996;11:1209-13.
64. Smyth CD, Miró F, Howles CM, Hillier SG. Effect of luteinizing hormone on follicle stimulating hormone-activated paracrine signalling in rat ovary. Hum Reprod 1995;10:33-9.
65. Cortvrindt R, Hu Y, Smitz J. Recombinant luteinizing hormone as a survival and diferentiation factor increases oocyte maturation in recombinant follicle stimulating hormone-supplemented mouse preantral follicle culture. Hum Reprod 1998;13:1292-302.
66. Fleming R, Lloyd F, Hebert M, Fenwick J, Griffiths T, Murdoch A. Effects of profound suppression of luteinizing hormone during ovarian stimulation on follicular activity, oocyte and embryo function in cycles stimlated with purified follicle stimulating hormone. Hum Reprod 1998;13:1788-92.
67. Porchet HC, Le Cotonnec JY, Neuteboom B, Canali S, Zanolo G. Pharmacokinetics of recombinant human luteinizing hormone after intravenous, intramuscular, and subcutaneous administration in monkeys and comparison with intravenous administration of pituitary human luteinizing hormone. J Clin Endocrinol Metab 1995;80:667-73.
68. Chandrasekher YA, Hutchinson JS, Zelinski-Wooten MB, Hess DL, Wolf DP, Stouffer RL. Initiation of periovulatory events in primate follicles using recombinant and native human luteinizing hormone to mimic the midcycle gonadotropin surge. J Clin Endocrinol Metab 1994;79:298-306.
69. Zelinski-Wooten MB, Hutchison JS, Hess DL, Wolf DP, Stouffer RL. Follicle stimulating hormone alones supports follicle growth and oocyte development in gonadotrophin releasing hormone antagonist-treated monkeys. Hum Reprod 1995;10:1658-66.
70. Hull M, Corrigan E, Piazzi A, Luomaye E. Recombinant human luteinizing hormone: an effective new gonadotropin preparation. Lancet 1994;344:334-5.
71. Kousta E, White DM, Piazzi A, Loumaye E, Franks S. Successful induction of ovulation and completed pregnancy using recombinant luteinizing hormone and follicle stimulating hormone in a woman with Kallmann´s syndrome. Hum Reprod 1996;11:70-1.
72. Agrawal R, West C, Conway GS, Page ML, Jacobs HS. Pregnancy after treatment with three recombinant gonadotropins. Lancet 1997;349:29-30.
73. The European Recombinant Human LH Study Group. Recombinant human luteinizing hormone (LH) to support recombinant human follicle-stimulating hormone (FSH)-induced follicular development in LH- and FSH-deficient anovulatory women: a dose-finding study. J Clin Endocrinol Metab 1998;83:1507-1514.
74. Zelinski-Wooten MB, Hess DL, Baughman WL y cols. Administration of an aromatase inhibitor during the late follicular phase of gonadotropin-treated cycles in rhesus monkeys: effects on follicle development, oocyte maturation, and subsequent luteal function. J Clin Endocrinol Metab 1993;76:988-95.
75. Wu TJ, Wang L, Wan YY. Detection of estrogen receptor messenger ribonucleic acid in human oocytes and cumulus-oocyte complexes using reverse transcriptase-polymerase chain reaction. Fertil Steril 1993;59:54-9.
76. Mannaerts B, Uilenbroek J, Schot P y cols. Folliculogenesis in hypophysectomized rats after treatment with recombinant human follicle-stimulating hormone. Biol Reprod 1994;51:72-81.
77. Weston AM, Zelinski-Wooten MB, Hutchison JS y cols. Developmental potential of embryos produced by in-vitro fertilization from gonadotrophin-releasing hormone antagonist-treated macaques stimulated with recombinant human follicle stimulating hormone alone or in combination with luteinizing hormone. Hum Reprod 1996;11:608-13.
78. Balasch J, Arroyo V, Carmona F y cols. Severe ovarian hyperstimulation syndrome: role of peripheral vasodilation. Fertil Steril 1991;56:1077-83.
79. Balasch J, Arroyo V, Fábregues F y cols. Neurohormonal and hemodynamic changes in severe cases of the ovarian hyperstimulation syndrome. Ann Inter Med 1994;121:27-33.
80. Balasch J, Arroyo V, Fábregues F, Jiménez W, Saló J, Vanrell JA. Immunoreactive endothelin plasma levels in severe ovarian hyperstimulation syndrome. Fertil Steril 1995a;64:65-8.
81. Manau D, Balasch J, Arroyo V y cols. Circulatory dysfunction in asymptomatic in vitro fertilization patients Relationship with hyperestrogenemia and activity of endogenous vasodilators. J Clin Endocrinol Metab 1998;83:1489-93.
82. Editorial. Ovarian hyperstimulation syndrome. Lancet 1991;338: 1111-2.
83. Navot D, Bergh PA, Laufer N. Ovarian hyperstimulation syndrome in novel reproductive technologies: prevention and treatment. Fertil Steril 1992;58:249-61.
84. Dourron NE, Williams DB. Prevention and treatment of ovarian hyperstimulation syndrome. Sem Reprod Endocrinol 1996;14:355-65.
85. Peinado JA, Molina I, Pla M y cols. Recombinant-human luteinizing hormone as ovulatory stimulus in superovulated does. J Assist Reprod Genet 1995;12:61-4.
86. Zegers-Hochschild F, Fernández E, MacKenna A y cols. The empty follicle syndrome: a pharmaceutical industry syndrome. Hum Reprod 1995;10:2262-5.