







El reconocimiento de subpoblaciones celulares reguladoras ha marcado una nueva etapa de desarrollo investigativo sobre la generación y control de la respuesta inmune. La subpoblación mejor caracterizada son los linfocitos T CD4+CD25+FOXP3+.
Uno de los grandes aportes a la comprensión de la regulación inmune es la identificación de FOXP3, un factor regulador de la transcripción, que participa directamente en la función de las células reguladoras T CD4+ humanas y murinas. FOXP3 ha sido definido por diversos autores como el gen maestro controlador del desarrollo y función de las células reguladoras y es considerado el principal marcador molecular, a la fecha, de esta subpoblación reguladora.
FOXP3 fue descrito inicialmente en ratones “escurfi”, los cuales presentan mutaciones espontáneas en el marco de lectura del gen denominado Foxp3. Este gen codifica para la proteína escurfina, cuya deficiencia está asociada a trastornos autoinmunes y linfoproliferativos severos. En los humanos, fue descrito el síndrome IPEX (inmunodisregulación poliendocrinopatía y endocrinopatía ligada al X), similar al de los ratones “escurfi” (2).
La investigación actual de la inmunología de células reguladoras apunta a la modulación de las respuestas mediadas por linfocitos T a través de la manipulación de la expresión de Foxp3 en modelos animales, lo que podría facilitar en el futuro la inducción de tolerancia a aloinjertos y el control de la autoinmunidad en la práctica clínica. En esta revisión se presentan los aspectos más importantes de la función de FOXP3 en la regulación inmune.
The recognition of regulatory cell subpopulations has marked a new stage of development on the research of the generation and control of the immune response. The best characterized subpopulation of T lymphocytes are the CD4+CD25+ cells; in this population, FOXP3+ has been identified as an associated phenotype, as this marker is not exclusive for this subpopulation.
One of the great contributions to the understanding of immune regulation is the identification of FOXP3, a regulatory transcription factor directly involved in the function of human and mouse CD4+ T regulatory cells. FOXP3 has been defined by various authors as the master gene controlling the development and function of regulatory cells and is considered the main marker for this regulatory subpopulation.
FOXP3 was initially described in “scurfy” mice, which have spontaneous mutations in the reading frame of Foxp3. This gene encodes the protein scurfin, whose deficiency is associated with autoimmune and lymphoproliferative disorders. In humans, the IPEX syndrome has been described (immunodysregulation polyendocrinopathy enteropathy X-linked syndrome), similar to that of scurfy mice.
The aim of the current research in the immunology of regulatory cells is to facilitate the future induction of allograft tolerance and control of autoimmunity in the clinical practice through the manipulation of FOXP3 expression. In this review, we discuss the most important aspects of the role of FOXP3 in immune regulation.
El nombre del dominio "forkhead" (FKH) se deriva del producto del gen forkhead (fkh) en Drosophila melanogaster, indispensable para la diferenciación terminal del embrión; la estructura de este dominio FKH fue identificada posteriormente en un grupo de factores de transcripción hepáticos con dominios de unión al DNA, hallazgo que dio lugar a la conformación del grupo de factores de transcripción con dominios FKH, los cuales están presentes en diversos organismos, desde levaduras hasta humanos. Muchos de los factores de este grupo están relacionados con la diferenciación embrionaria(1-3).
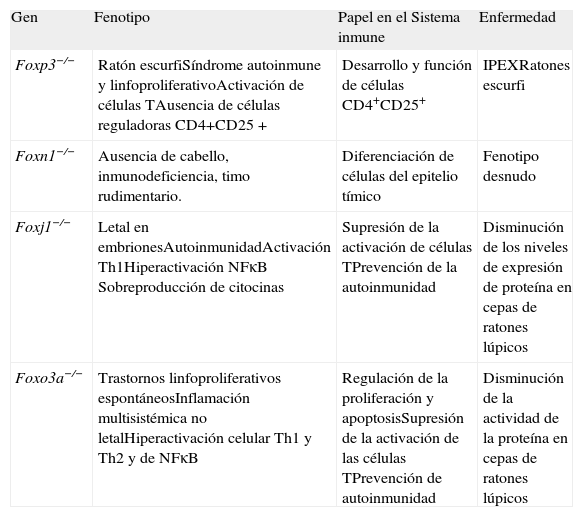
Bajo la denominación de FOX se designan los factores de transcripción con dominio "forkhead" en los vertebrados, clasificados con base en su estructura. Los factores de transcripción FOX con acción en el sistema inmune son FOXJ1, FOXN1, FOXO3A y FOXP3. En la Tabla I se resumen las características de estos factores de transcripción y el fenotipo de los ratones knockout para estos genes(3).
Factores de transcripción Fox y fenotipos de los ratones knockout
| Gen | Fenotipo | Papel en el Sistema inmune | Enfermedad |
| Foxp3−/− | Ratón escurfiSíndrome autoinmune y linfoproliferativoActivación de células TAusencia de células reguladoras CD4+CD25+ | Desarrollo y función de células CD4+CD25+ | IPEXRatones escurfi |
| Foxn1−/− | Ausencia de cabello, inmunodeficiencia, timo rudimentario. | Diferenciación de células del epitelio tímico | Fenotipo desnudo |
| Foxj1−/− | Letal en embrionesAutoinmunidadActivación Th1Hiperactivación NFκB Sobreproducción de citocinas | Supresión de la activación de células TPrevención de la autoinmunidad | Disminución de los niveles de expresión de proteína en cepas de ratones lúpicos |
| Foxo3a−/− | Trastornos linfoproliferativos espontáneosInflamación multisistémica no letalHiperactivación celular Th1 y Th2 y de NFκB | Regulación de la proliferación y apoptosisSupresión de la activación de las células TPrevención de autoinmunidad | Disminución de la actividad de la proteína en cepas de ratones lúpicos |
Adaptado de (3).
La cristalografía del dominio FKH reveló la presencia de una estructura similar a la forma de una mariposa, conformada por dobles hélices aladas. Esta estructura es poco variable y contiene aminoácidos conservados entre los diferentes factores de transcripción. A diferencia de la homología en los dominios de unión al DNA, los factores de transcripción FOX presentan poca o ninguna homología en la secuencia de los dominios de activación o represión entre sí, y se conoce poco sobre su interacción con la maquinaria transcripcional(1,3).
Foxp3 pertenece a la subfamilia Foxp, que incluye a Foxp1, Foxp2, Foxp3 y Foxp4. Foxp1 ha sido implicado como un gen supresor de tumores, puesto que la pérdida de su función se asocia a trastornos linfoproliferativos, cáncer de pulmón, mama y estómago. Foxp2 ha sido implicado en trastornos del lenguaje y en el desarrollo neuronal, Foxp3 en la generación y función de las células reguladoras CD4+CD25+ y Foxp4 se ha asociado al desarrollo de la corteza cerebral anterior en modelos murinos(4,5).
Aunque todas las proteínas FOX se unen al DNA, algunas ejercen efectos transactivadores y otras, efectos inhibidores de la transcripción. El dominio FKH de FOXP3 está localizado muy cerca del extremo carboxiterminal de la proteína, lo que sugiere que carece de dominio transactivador y lleva a postular que podría actuar como represor transcripcional(3).
Entre las evidencias para proponer que Foxp3 actúa principalmente como inhibidor de la transcripción se encuentra que, primero, las células T de los ratones "escurfi" (deficientes de Foxp3) presentan un estado constante de hiperactivación e hiperrespuesta a la estimulación a través del TCR (receptor de las células T). Segundo, las células T de los ratones transgénicos para Foxp3 presentan pobre respuesta a la estimulación del TCR, proliferan poco y producen bajas cantidades de IL-2 (interleucina 2) y tercero, la sobreexpresión de FOXP3 en células Jurkat, una línea de células T de leucemia humana, inhibe la transcripción de IL-2 por mecanismos posiblemente competitivos, debido a que el sitio de unión de Foxp3 en el promotor de IL-2 se sobrepone al de NFAT (factor activador de células T), uno de los factores de transcripción más importantes para IL-2(4,6).
Otros efectos de inhibición transcripcional fueron reportados por Betelly y cols., quienes describieron que Foxp3 suprime la producción de IL-2, IL-4 e IFN-γ (interferón γ) por mecanismos que incluyen asociación a los factores de transcripción de la familia Rel, NF-κB y NFAT y bloqueo de su unión a los promotores de las citocinas mencionadas. Por otro lado, se identificaron sitios potenciales de unión de Foxp3 en los promotores de los genes que codifican IL- 4, TNF-α (factor de necrosis tumoral α) y GM-CSF (factor estimulante de colonia granulocito-monocito), citocinas cuya regulación está alterada en los ratones "escurfi"; sin embargo, las consecuencias funcionales de estas observaciones no han sido completamente determinadas(4).
Además de lo anterior, también se ha propuesto que Foxp3 podría ejercer efectos transactivadores. La transfección de Foxp3 a células CD4+CD25- se asocia con la adquisición de la función y el fenotipo de las células reguladoras CD4+CD25+, que incluye, CTLA-4high (Antígeno del linfocito T citotóxico), GITRhigh (Receptor del factor de necrosis tumoral inducido por glucocorticoides) y CD25high; se postula que Foxp3 podría inducir la transcripción de los genes de estas moléculas, pero aún se desconoce cuáles son los mecanismos(7).
Los efectos que tiene FOXP3 sobre la inhibición o activación de algunos genes con importancia en la regulación inmune se resumen en la Tabla II.
IDENTIFICACIÓN DE FOXP3Foxp3 fue identificado inicialmente en los ratones "escurfi". Estos ratones poseen una mutación espontánea ligada al cromosoma X, descrita por primera vez por Russell y cols. en 1959; los machos afectados presentan descamación cutánea, pérdida de peso, diarrea, mal absorción, linfadenopatías, hepatoesplenomegalia e infiltrado de linfocitos en diferentes órganos, que conducen a la muerte entre la tercera y cuarta semana de vida. Estos hallazgos son similares a los presentados en ratones deficientes de CTLA-4 o TGF-β (Factor transformante de crecimiento β), los cuales fallecen a la tercera semana de vida y presentan unos recuentos de leucocitos en sangre periférica 20 veces mayores de lo normal(1,8).
Posteriormente, en 1991, Godfrey y cols. sugirieron que los ratones "escurfi" podrían ser el resultado de un tipo de disregulación inmune y no de una inmunodeficiencia como las reconocidas hasta la fecha. Observaron que el timo de los ratones jóvenes, aunque de menor tamaño, se encontraba densamente poblado de linfocitos y con clara diferenciación corticomedular; sin embargo, a medida que la enfermedad progresaba, la corteza tímica era rápidamente depletada, mientras que los órganos linfoides periféricos estaban marcadamente infiltrados con signos de hematopoyesis extramedular. Finalmente, los ratones desarrollaban enfermedades linfoproliferativas mediadas por linfocitos T CD4+CD8-(1,9,10).
Entre las alteraciones inmunológicas de los ratones "escurfi" se describe hiperactivación de las células T, asociada a la expresión de CD69, CD25, CD80 y CD86. Las células T CD4+ de estos ratones producen 1000 veces más GM-CSF que las células CD4+ de ratones silvestres, y producen mayor cantidad de otras citocinas como IL-2, IL-5, IL-6, IL-7, IL- 10, IFN-γ y TNF-α. El estado de hiperrespuesta es resistente a la supresión por inhibidores de las tirosina-cinasas como genisteína y herbimicina y a inmunosupresores como la ciclosporina A, lo que sugiere que la regulación de la activación de la célula T está alterada. Se observa además un aumento de monocitos Mac-1+ y disminución de células B220+ en órganos linfoides secundarios(1,11).
Sólo en el 2001 Brunkow y cols. reportaron una mutación por inserción de 2 pares de bases que altera el marco de lectura del gen que codifica para un regulador transcripcional de la familia "Forkhead", el cual es el responsable de las alteraciones inmunológicas observadas en los ratones "escurfi"; como resultado de la mutación se traduce una proteína funcionalmente inactiva que carece del dominio de unión al DNA. Estos autores identificaron este gen por mapeo físico y análisis de secuencia, y lo denominaron Foxp3. El locus de la mutación "escurfi" fue mapeado en la región proximal del cromosoma X, a un intervalo de 1.7 CM de DXWas70 y Otc(3,12).
Actualmente es bien sabido que los ratones "escurfi" carecen de células reguladoras CD4+CD25+FOXP3+ y presentan un mayor número de células efectoras CD4+CD25+ por activación celular crónica. Igualmente se conoce que la escurfina murina está conformada por 429 aminoácidos y además del dominio FKH, tiene un dominio de prolina, un dedo de zinc C2H2 y un motivo de cierre de leucina (Figura 1). Su expresión se observa especialmente en células CD4+CD25+(3).
.")
El gen FOXP3 humano está localizado en la región cromosómica Xp11.23. Está formado por 11 exones que codifican una proteína de 431 aminoácidos, la cual presenta un 86% de homología con la murina. El gen es expresado principalmente en tejidos linfoides como timo, bazo y ganglios linfáticos, en células CD4+CD25+ y en menor proporción en células CD4+CD25−, células CD4+CD8+, células CD8+ y timocitos CD4-CD8-αβ TCRhi(11).
Hasta el momento, han sido descritas dos principales isoformas de FOXP3 en los humanos, FOXP3 y FOXP3Δ2 (que carece del exón 2). Para evaluar el papel de estas dos isoformas, Allan y cols. emplearon un modelo de sobreexpresión de ambas isoformas basado en transducción de vectores retrovirales. Observaron que la transducción de células T CD4+CD25− con una de las dos isoformas induce un estado de hiporespuesta y disminución de la producción de IL-2; sin embargo, sólo cuando son transducidas las dos isoformas, se induce el fenotipo y la función reguladora. Lo que sugiere que ambas isoformas actúan de forma conjunta a nivel molecular para influir en la transcripción de genes asociados a las células reguladoras CD4+CD25+(13).
DE LOS RATONES "ESCURFI" AL SÍNDROME IPEXLa deficiencia de escurfina por mutaciones en el gen se asocia a trastornos autoinmunes y linfoproliferativos más severos que la ausencia de las células T CD4+CD25+, por lo que se postula que su acción en la regulación inmune es más amplia. En humanos, la enfermedad relacionada con la deficiencia de FOXP3 es IPEX, o síndrome de disregulación inmune poliendrocrinopatía y enteropatía ligado al X, descrito en 1982 en una familia con 19 hombres afectados en cinco generaciones. IPEX afecta a los hijos varones de madres portadoras de la mutación de FOXP3, los cuales presentan enfermedades como diabetes, anemia hemolítica, tiroiditis, enfermedad linfoproliferativa masiva, alergias, eczema, enfermedad inflamatoria intestinal, o dermatitis exfoliativa, entre otras. Los afectados por esta enfermedad fallecen en los primeros dos años de vida por alteraciones metabólicas y sepsis(9,12,14).
No es claro si los pacientes presentan mayor susceptibilidad a las infecciones por la alteración genética, a consecuencia de la alteración de la función de las barreras de la piel y el tracto gastrointestinal, o debido a los inmunosupresores administrados en el tratamiento. Los patógenos más comúnmente encontrados en los pacientes con IPEX son Enterococcus, Staphylococcus, Cytomegalovirus y Candida los cuales llevan a desarrollar sepsis, meningitis, neumonía y osteomielitis(11,14).
Las mujeres portadoras no presentan fenómenos autoinmunes, pero un gran número de sus células reguladoras presentan la función alterada. En este caso, la autotolerancia es mantenida a expensas de las células reguladoras cuyo cromosoma X sin la mutación no sufre lionización(11).
En humanos, se han reportado más de 20 mutaciones diferentes, especialmente mutaciones sustitutivas en la porción C-terminal del dominio de unión al DNA. Otras mutaciones descritas afectan la región rica en prolina y el cierre de leucina de la porción N-terminal de la proteína. Entre el 10 y el 40% de pacientes presentan el fenotipo de IPEX sin mutación en FOXP3, lo que sugiere la posible existencia de alteraciones en las regiones promotoras, en los potenciadores, o en otros genes que se relacionan directa o indirectamente con la activación de FOXP3(11,12).
El estudio de varias familias ha revelado cambios de nucleótidos únicos en el dominio "forkhead" de FOXP3, alterando su unión al DNA. Otras alteraciones descritas son la deleción de tres pares de bases en el cierre de leucina del dominio de dimerización, alterando la homodimerización y heterodimerización, afectando así la función de FOXP3(1,11,12,14,15).
Para el tratamiento del IPEX se han empleado inmunosupresores como Ciclosporina A y FK-506 (tacrolimus), solos o en combinación con esteroides, sin embargo, la respuesta no es satisfactoria. En la actualidad, el único tratamiento que se considera curativo es el trasplante de médula ósea. Un futuro tratamiento podría ser la reconstitución de los pacientes con células autólogas CD4+CD25+ transducidas con el gen FOXP3(11,12).
POLIMORFISMOS DEFOXP3EN OTRAS ENFERMEDADESLa evaluación de mutaciones y polimorfismos de FOXP3 es de interés, no solo en trastornos como IPEX, sino en otras enfermedades autoinmunes, en las que se ha evaluado la participación de las células reguladoras. Park y cols. evaluaron 10 polimorfismos de nucleótidos únicos en un grupo de controles sanos y dos grupos de pacientes, el primero con cirrosis biliar primaria y el segundo con enfermedad de Crohn. En esta enfermedad inflamatoria intestinal se ha reportado, en modelos animales, que la transferencia adoptiva de células reguladoras CD4+CD25+ controla la inflamación y restablece la arquitectura normal de la mucosa intestinal. Estos autores encontraron que el SNP (polimorfismo de un solo nucleótido) IVS9+459 en FOXP3 está asociado significativamente a cirrosis biliar, pero no encontraron SNPs asociados a enfermedad de Crohn(16).
En esclerosis múltiple, una enfermedad neurológica compleja y devastadora, Huan y cols. reportaron la disminución de la expresión del mRNA y la proteína FOXP3 en las células reguladoras CD4+CD25+, indicando que alteraciones en la inmunoregulación contribuyen a la fisiopatología y postulando la posibilidad de utilizar terapias basadas en la inducción de FOXP3 como posible tratamiento(6,17).
FOXP3, GEN CONTROLADOR MAESTRO DEL DESARROLLO Y FUNCIÓN DE LAS CÉLULAS REGULADORAS CD4+CD25+La semejanza entre las alteraciones inmunológicas asociadas a mutaciones en Foxp3 y la depleción de las células reguladoras CD4+CD25+, motivó la investigación de la relación entre este gen y el desarrollo y la función de las células reguladoras. Experimentos en ratones han demostrado que el mRNA de Foxp3 y la proteína escurfina se expresan específicamente en las células reguladoras CD4+CD25+, mientras que los marcadores de superficie CD25, CTLA-4, GITR, y CD103 pueden estar presentes luego de la activación celular en células sin función reguladora(15).
Una de las aproximaciones empleadas para evaluar la acción de Foxp3 en la regulación inmune ha sido la generación de ratones transgénicos, los cuales han permitido determinar los efectos de la sobreexpresión de Foxp3 in vivo. Brunkow y cols. generaron cepas de ratones con diferentes niveles de expresión del transgen de foxp3. Cada una de estas cepas, cuando fue cruzada con la cepa salvaje, presentó disminución de la celularidad en los órganos linfoides, proporcional a la expresión del transgen de Foxp3. A mayor expresión del transgen, mayor era la disminución de la celularidad en el bazo a expensas de las poblaciones de linfocitos CD4+ y CD8+, mientras que la celularidad del timo se conservaba. La cepa con una expresión de Foxp3 tres veces mayor de lo normal presentó 50% de disminución de células CD4+ y 75% de disminución de células CD8+. Las células CD4+ de estos ratones, al ser estimuladas in vitro mostraron una actividad proliferativa disminuida y no produjeron IL-2(18).
Sin embargo, sólo hasta el reporte de Hori y cols. se comprobó que Foxp3 está relacionado directamente con generación de la función reguladora. En experimentos en los que células CD4+CD25- fueron transducidas con un vector retroviral que portaba el gen de Foxp3, estos autores observaron que las células transducidas adquirían la función y el fenotipo de superficie de las células reguladoras naturales. Luego de la transducción, las células presentaban alta expresión de CD25, CD103, CTLA-4, GITR y mRNA de IL-10 e inhibían la proliferación de células T estimuladas con anti-CD3 más anti-CD28(2,6).
De manera similar a lo observado en el modelo murino, FOXP3 en humanos participa en la generación y función de las células reguladoras CD4+CD25high. La expresión del mRNA y de la proteína FOXP3 es alta en las células reguladoras CD4+CD25high y 100 veces menor en las células CD4+CD25-, las cuales tienen la capacidad de convertirse en células reguladoras adaptativas. FOXP3 también se expresa en un bajo porcentaje de células reguladoras CD8+(5,7).
En un modelo murino de enfermedad injerto contra huésped (EICH), Albert y cols. demostraron que la transducción de un clon de células CD4+ Th1 antígeno específicas con Foxp3 permite obtener células T reguladoras antígeno específicas con capacidad de suprimir la respuesta alogénica in vitro y prevenir la EICH in vivo, de igual forma que las células reguladoras naturales CD4+CD25+. Estos hallazgos aportan evidencias de la función de Foxp3 in vivo y en un futuro podría ser posible el desarrollo de estrategias terapéuticas basadas en transducción de Foxp3 aplicables a los trasplantes alogénicos(19).
Es importante resaltar que aunque Foxp3 ha sido ampliamente asociado a la generación y función de las células reguladoras, es posible que este gen tenga acciones más amplias en la regulación inmune, puesto que la deficiencia de escurfina produce trastornos autoinmunes y linfoproliferativos más severos que los observados en ausencia de las células T CD4+CD25+(20).
La función de Foxp3 en células no T ha sido muy poco estudiada; entre los reportes al respecto Chang y cols., en el 2005, describen alteraciones en la timopoyesis en los ratones "escurfi" relacionadas con la inactivación de Foxp3 en el estroma tímico y la sobreexpresión de ErbB2 en las células estromales. Las descripciones iniciales de los efectos de la mutación de Foxp3 en células no-T, abren un campo investigativo que podría ampliar el conocimiento existente acerca de otros mecanismos involucrados en la regulación inmune(21).
FACTORES RELACIONADOS CON LA EXPRESIÓN DE FOXP3Aunque se han identificado varias vías de señalización que inducen la expresión de FOXP3, los mecanismos precisos por los cuales la expresión de esta proteína es controlada en las células Tregs no están bien entendidos. Acciones sinérgicas de señalización a través del TCR, moléculas coestimuladoras y receptores de citocinas son requeridas para inducir la transcripción de FOXP3 (Figura 2)(22).
 (interleucinas 2,4,7,9,15, y 21) y el receptor del factor estimulante de crecimiento β (TGF-βR). Estos eventos resultan en la activación de factores de transcripción que participan en la expresión de FOXP3, incluyendo CREB, ATF, SP1, NFAT, AP1, TIEG1, SMAD3, y STAT5. Sin embargo, la contribución de cada vía de señalización (para inducir o reprimir la activación) en la expresión de FOXP3 puede diferir entre los diferentes tipos de células. Adaptado de (22).")
Diferenciación de células T reguladoras. La expresión de Foxp3 es dependiente de la señalización a través del TCR, del CD28 y de receptores de citocinas que comparten la cadena gamma común γ (γc) (interleucinas 2,4,7,9,15, y 21) y el receptor del factor estimulante de crecimiento β (TGF-βR). Estos eventos resultan en la activación de factores de transcripción que participan en la expresión de FOXP3, incluyendo CREB, ATF, SP1, NFAT, AP1, TIEG1, SMAD3, y STAT5. Sin embargo, la contribución de cada vía de señalización (para inducir o reprimir la activación) en la expresión de FOXP3 puede diferir entre los diferentes tipos de células. Adaptado de (22).
En humanos, la activación a través del TCR induce la unión del NFAT y de la proteína activadora 1 (AP1) al promotor de FOXP3. En ratones, la activación por el TCR resulta en la unión del elemento de respuesta al AMP cíclico (CREB) o del factor de activador de transcripción (ATF) a un elemento potenciador en el gen de Foxp3. Posterior a la activación de las células T por el TCR, NFAT forma un complejo con AP1, e inducen la transcripción de genes asociados con activación. En condiciones de activación parcial, NFAT activa un grupo de genes independientes de AP1, los cuales codifican reguladores negativos de la activación de las células T, llevando a un estado de anergia(23). Yongqing Wu y cols. demostraron que FOXP3 tiene la capacidad de formar un complejo con NFAT y el DNA (NFAT-FOXP3- DNA) que se asemeja al complejo formado por NFAT-AP1- DNA, indicando que el sistema inmune emplea NFAT como un regulador común tanto para la activación de células T efectoras (NFAT-AP1) como para la inducción de células Treg (NFAT-FOXP3)(23,24).
Otra de las proteínas que se han visto involucradas en la activación de FOXP3 a través de la señalización por el TCR es la proteína kinasa C- (PKC-), sugiriendo que PKC- promueve el desarrollo de Tregs potenciando la expresión de FOXP3 mediante la activación de la vía de Ca2+− Calmodulina-NFAT; se ha observado además que la participación de moléculas coestimuladoras como CD28 podría estar involucrada también en la inducción del fenotipo regulador(23,24).
La señalización a través de diversos receptores de citocinas que comparten la cadena común γ (γc), como es el caso del receptor de IL-2, se ha visto involucrada en la inducción de la expresión de FOXP3, mediante la activación de diferentes vías de señalización que involucra las proteínas JAK/STAT y el receptor de TGF-β que involucra proteínas de la familia SMAD(22,23).
La identificación de moléculas o condiciones de estimulación celular que controlan la expresión de FOXP3 es importante, no solo para entender mejor la biología de la regulación inmune, sino también para el desarrollo de aproximaciones terapéuticas en trasplantes, autoinmunidad, alergia y cáncer. Por esta razón discutiremos un poco más sobre cada uno de estos factores.
ESTÍMULOS A TRAVÉS DEL TCR- ACTIVACIÓN CELULARWalker y cols. demostraron que la expresión de FOXP3 es inducible en células humanas al estimular células mononucleares de sangre periférica con anti-CD3 más anti- CD28. Estos autores reportaron que además de la expresión de CD25 y la producción de citocinas, la estimulación a través del TCR induce la expresión de FOXP3 en células que no tenían actividad reguladora previa; las células CD4+CD25highFOXP3+ generadas por la activación adquieren la función reguladora y son capaces de disminuir la proliferación de las células respondedoras CD4+CD25−, de forma similar a las células reguladoras naturales, a través de mecanismos dependientes de contacto célula-célula e independiente de citocinas. La expresión de FOXP3 se observó a las 24 horas del cultivo, con un pico máximo de expresión a las 72 horas, asociada a la expresión de CD25 en la superficie. Estos autores no observaron la inducción de Foxp3 en células murinas, bajo las mismas condiciones experimentales. Estas evidencias sugieren que la generación de células reguladoras de novo es una consecuencia natural de la respuesta inmune en humanos y la falla en su generación puede contribuir a enfermedades autoinmunes(15).
EL FACTOR TRANSFORMANTE DE CRECIMIENTO β, TGF-β, INDUCE LA EXPRESIÓN DE FOXP3Una de las principales citocinas relacionadas con inducción de células reguladoras CD4+CD25+ es TGF-β . En un modelo in vitro con células murinas, Chen y cols. demostraron que TGF-β convierte a las células CD4+CD25− en células reguladoras CD25+, cuando han sido estimuladas a través de su TCR con anti-CD3 y un tercer estímulo como anti-CD28 o células presentadoras de antígeno. Las células reguladoras generadas por la estimulación con TGF-β expresan FOXP3 dependiente de la dosis y presentan un fenotipo CD25high, CTLA-4 intracelularhigh, TGF-β de membrana y CD45RB-/low, similar al de las células reguladoras naturales. Estos autores demostraron que en las condiciones de estimulación descritas, TGF-β induce la expresión de FOXP3 de novo en las células CD25−, un efecto no observado por la estimulación con IL- 10. Las células reguladoras generadas ejercen una función supresora in vitro dependiente del contacto célula a célula, e igualmente son supresoras in vivo en un modelo murino de asma inducida por aero alérgenos. En estos experimentos, TGF-β no incrementa la expresión de FOXP3 en las células reguladoras naturales que constitutivamente expresan altas cantidades de FOXP3; sin embargo, existen evidencias controversiales al respecto(25).
Fantini y cols. describieron que el estímulo con TGF- β1 más anti-CD3 y anti-CD28 induce la expresión de FOXP3 en células CD4+CD25− murinas y humanas de sangre periférica con un fenotipo y una función reguladora(26).
Modelos in vivo de trasplante cardíaco murino, demostraron que la transfección de TGF-β 1 humano en las células del órgano trasplantado, prolonga la sobrevida del injerto por mecanismos que incluyen la inducción de la expresión de Foxp3 y la generación de células Tregs en el receptor. La transferencia adoptiva de esplenocitos de los ratones que recibieron los aloinjertos transfectados con TGF-β1 induce tolerancia en ratones trasplantados con órganos no transfectados con TGF-β1, lo que se relaciona con la generación de mecanismos de tolerancia mediados por células reguladoras(27).
LA INTERLEUCINA 2 ES ESENCIAL PARA EL MANTENIMIENTO DE LA FUNCIÓN REGULADORA EN PERIFERIALa mayoría de las células reguladoras T CD4+ expresan constitutivamente altos niveles de la cadena alfa del receptor de IL-2, CD25; sin embargo, no se conoce completamente la función de IL-2 en la biología de las células reguladoras. Para evaluar el papel de la IL-2, Fontenot y cols. emplearon ratones knock-in para Foxp3 (gpf) y deficientes para IL-2 (IL-2−/−) o para el IL-2Rα (IL-2Ra−/−). Estos autores reportaron que la IL-2 no es indispensable para la inducción de la expresión de Foxp3 en el timo y para la acción reguladora in vitro; sin embargo, en ensayos in vivo, la IL-2 es indispensable para el mantenimiento de la expresión de Foxp3 y la función de las células Tregs en la periferia. También observaron que Foxp3 no se expresa en el timo ni en la periferia de ratones deficientes de la cadena gama del receptor de IL-2, IL-2Rγ, la cual forma parte de los receptores de IL-15 e IL-4, lo que sugiere la participación de otras citocinas diferentes a IL-2 en la expresión tímica de FOXP3(28).
LOS ESTRÓGENOS INDUCEN LA EXPRESIÓN DE FOXP3Polanczyk y cols. reportaron que el 17-β estradiol, E2, aumenta la expresión de Foxp3 in vivo e in vitro. El tratamiento de células murinas con E2 incrementó el número de células CD25+ y el nivel de expresión de Foxp3 y protegió a los animales de desarrollar encefalomielitis autoinmune experimental, esta misma protección fue observada durante el proceso de gestación en ratones. Los hallazgos de este estudio podrían sugerir que la inducción de Foxp3 y, a su vez, de células Tregs, podría ser uno de los mecanismos de tolerancia materno-fetal inducido por estrógenos(29).
PROSTAGLANDINA E2 ACTUA COMO INMUNOSUPRESOR AL INDUCIR FOXP3 EN MODELOS DE CÁNCERBaratelli y cols. reportaron que la prostaglandina E2 (PGE2), un producto de la ciclooxigenasa, aumenta la función reguladora in vitro de las células Tregs humanas e induce un fenotipo regulador en las células CD4+CD25−, por mecanismos que incluyen la inducción e incremento del mRNA y la expresión de la proteína FOXP3(30).
Por otro lado, Sharma y cols. demostraron que la PGE2, actúa como inmunosupresor y contribuye a la generación de carcinoma de células pequeñas de pulmón incrementando la expresión de FOXP3 en las células reguladoras CD4+CD25+. El uso de inhibidores de la ciclooxigenasa-2 suprime la acción de las células reguladoras y mejora la respuesta antitumoral. Esto indica que sobreexpresión de FOXP3 tiene efectos deletéreos, siendo la inhibición de FOXP3 un blanco terapéutico en algunas patologías tumorales(31).
REGULACIÓN EPIGENÉTICA DE FOXP3La regulación epigenética es crucial para controlar la expresión del locus de FOXP3. Acetilaciones y metilaciones que ocurren en sitios específicos en proteínas histonas, son esenciales para la remodelación de la cromatina, permitiendo o no el acceso de factores de transcripción hasta la región promotora del gen. Adicionalmente, los motivos CpG ubicados dentro de las regiones promotoras, también pueden ser blanco de metilaciones o demetilaciones, llevando a la condensación o descondensación de la cromatina respectivamente, afectando así la expresión génica(22).
En ratones y humanos, se han encontrado diferentes patrones de metilación en el locus de FOXP3 que difiere entre las Tregs y T convencionales (Tconv). Análisis de secuencia han revelado tres regiones altamente conservadas en el locus de FOXP3, involucradas con regulación de la transcripción. La primera región, es la región promotora para FOXP3, localizada 6.5 kb corriente arriba del primer exón que codifica para FOXP3. Esta región contiene motivos CpG, los cuales se encuentran demetilados en Tregs, mientras que están débilmente metilados en las Tconv. Adicionalmente, el promotor de FOXP3 muestra una mayor asociación con histonas acetiladas en las Tregs que en las Tconv, lo que sugiere que este promotor se encuentra más asequible en la población reguladora (Figura 4)(22).
.")
Origen de células Tregs FOXP3+ en el timo. Las células T reguladoras en el timo se generan a partir de un precursor de linfocitos T CD4+ en un proceso que no está completamente dilucidado, pero se conoce que las células del epitelio cortical y medular participan de forma activa en dicha generación. También se ha propuesto que citocinas γc participan en la diferenciación a Tregs. Adaptado de (22).
.")
Control epigenético sobre el locus de FOXP3. Se han identificado tres regiones no codificantes las cuales están involucradas con el control epigenético de FOXP3. Modificaciones epigenéticas en estas tres regiones, que incluyen acetilación de histonas y metilación del DNA son descritas para células Tconv, células Treg inducibles por TGF-β y células Treg naturales. Adaptado de (22).
La segunda región no codificante altamente conservada, ha sido identificada como un elemento sensible al TGF-β que contiene sitios de unión para NFAT y SMADs. El alto número de histonas H4 acetiladas en esta región, permite que el DNA se encuentre más asequible, efecto observado tanto en las Tregs naturales como en las Tregs inducibles por TGF-β (Figura 4)(22).
La tercera región es definida por Huehn y cols. como región demetilada específica de células Tregs (TSDR), la cual se encuentra totalmente demetilada en Tregs y metilada en células Tconv. Recientemente, se ha postulado esta región no como la encargada del encendido y el apagado de la transcripción de FOXP3, sino como una región determinante de la estabilidad de la expresión de dicha proteína. TSDR demetilada se relaciona con expresión estable de FOXP3 (células Treg naturales), mientras que la expresión transitoria de FOXP3 (células Treg inducibles y Tconv) se asocia con metilación de la región TSDR (Figura 4)(22).
FACTORES TÍMICOS RELACIONADOS CON LA EXPRESIÓN DE FOXP3Actualmente se conoce poco acerca de los factores tímicos relacionados con la generación de un linaje regulador. Fontenot y cols., al evaluar la generación de timocitos Foxp3+ durante la ontogenia, demostraron que su desarrollo es más tardío con respecto al de los timocitos no reguladores. Estos autores postularon la posible existencia de señales generadas por la médula del timo que podrían favorecer el desarrollo de un linaje de células reguladoras, independiente de los timocitos no reguladores, CD4+ o CD8+(32).
Watanabe y cols. reportaron que las células dendríticas CD11c+ activadas con linfopoyetina estromal del timo (TSLP), producida por los corpúsculos de Hassall humanos, median la diferenciación de los timocitos CD4+CD8−CD25− a CD4+CD25+FOXP3+. En estudios de inmunohistoquímica, se observó la asociación de células CD25+CTLA-4+ con células dendríticas activadas y corpúsculos de Hassall-TSLP+, en la médula del timo. Este reporte es de especial interés debido a que describe cómo la médula tímica puede estar involucrada en la diferenciación de timocitos FOXP3+ y una función para los corpúsculos de Hassall, los cuales fueron descritos hace más de 50 años, pero cuya función aun no se conoce completamente(33).
En varios estudios se ha observado que para el desarrollo de células reguladoras en el timo es necesario el desarrollo de un TCR de alta afinidad por péptidos propios; estas células escapan a la regulación negativa por mecanismos que aún no han sido del todo clarificados (Figura 3).
Las células reguladoras naturales se generan en el timo a partir de linfocitos T CD4+ vírgenes que expresan un TCR de alta afinidad para el complejo MHC-péptido. Esta señal es necesaria para la inducción de la expresión de FOXP3(34). Este proceso es dependiente de la señalización por CD28, de citocinas que señalizan por receptores que comparten la cadena γc y otros factores no identificados. Hasta el momento, no se conocen los mecanismos por los cuales las células reguladoras escapan a la selección tímica(35,36).
EFECTOS DE LOS MEDICAMENTOS INMUNOSUPRESORES EN LA EXPRESIÓN DE FOXP3Baan y cols. evaluaron el efecto de los medicamentos inmunosupresores en la expresión de FOXP3 en el curso de la reacción alogénica en cultivos mixtos de linfocitos. Al quinto día del cultivo, el mRNA de FOXP3 se incrementa de 10 a 20 veces por encima del valor inicial, lo que sugiere que la generación de las células reguladoras es una consecuencia normal de la activación celular durante la reacción alogénica. La adición de Ciclosporina A, Tacrolimus, Daclizumab, o anticuerpos monoclonales anti-CD25, inhiben en un 70-90% la transcripción de FOXP3, efecto no observado al adicionar Rapamicina a los cultivos. El trabajo de estos autores coincide con evidencias previas que sugieren que inmunosupresores de uso frecuente en la actualidad, como la Ciclosporina A, impiden el establecimiento de mecanismos de tolerancia, mientras que otros fármacos actúan como inmunomoduladores sin alterar estos mecanismos(37).
APROXIMACIÓN A LA EVALUACIÓN DE FOXP3Teniendo en cuenta que FOXP3 es una proteína intracelular, no es fácil detectarla, lo que dificulta el estudio de la biología de este factor de transcripción en las células que lo expresan. La cuantificación del mRNA es una de las metodologías más usadas para evaluar FOXP3, sin embargo, los niveles de mRNA podrían no reflejar de forma fiable los niveles de proteína FOXP3 expresada. Muchas metodologías se han empleado para la evaluación de la expresión de FOXP3, entre ellas, Western-blot, RT-PCR, PCR en tiempo real, inmunohistoquímica de tejido y citometría de flujo, entre otras(38).
El desarrollo del modelo animal de Ratones FIR (Foxp3- IRES-mRFP), es de especial interés porque permite evaluar la biología de las células que expresan Foxp3. El modelo murino desarrollado por Wan y Flavell, en el cual un gen reportero bicistrónico que expresa una proteína roja fluorescente fue insertado en el locus endógeno Foxp3 en ratones con fondo genético B6, constituye un modelo de estudio interesante para evaluar la generación de células reguladoras Foxp3+ in vivo y podría ser muy útil como fuente de células para estudios de transferencia adoptiva(39).
CONCLUSIONESLa regulación de la respuesta inmune es uno de los principales objetivos de investigación de la inmunología actual. Desde perspectivas tan diversas como autoinmunidad, infección, cáncer y tolerancia a trasplantes, los mecanismos involucrados en la regulación inmune están siendo intensamente estudiados para contribuir, no solo al conocimiento de la biología del sistema inmune, sino para ampliar la comprensión de la fisiopatología de diversas enfermedades y el diseño de estrategias terapéuticas.
Si bien FOXP3 puede expresarse luego de la activación celular, esta expresión es transitoria y menor a la observada en las células T reguladoras naturales. El reconocimiento de FOXP3 como el gen controlador maestro del desarrollo y función de las células Tregs, es uno de los aportes más significativos a la comprensión de la regulación de la respuesta inmune. El conocimiento existente en el momento actual permite proyectar que en un futuro cercano podría ser posible diseñar estrategias de inmunointervención basadas en su inducción o represión, no solo en modelos experimentales, sino en el contexto clínico. En los próximos años, la investigación de los mecanismos moleculares de FOXP3 y otras moléculas, que muy seguramente serán descritas, resolverá muchas de las preguntas que nos hacemos sobre las funciones del sistema inmune, especialmente sobre la tolerancia.
AGRADECIMIENTOSTrabajo financiado por Colciencias (código111540820527).
CONFLICTO DE INTERÉSLos autores declaran no tener conflicto de interés.