






Introducción
La arteriosclerosis en humanos es un proceso de desarrollo lento y es la causa principal de las enfermedades cardiovasculares y cerebrovasculares, que causan una elevada mortalidad y morbilidad en los países desarrollados. La hiperlipidemia, la inflamación, la oxidación y la nutrición son factores clave en la aparición y progresión de la arteriosclerosis.
Debido a la dificultad de estudiar la arteriosclerosis en humanos, el ratón deficiente para la apolipoproteína E (apo E) ha sido muy utilizado como modelo gracias a que su metabolismo lipídico se parece mucho al de los humanos y a que el desarrollo de su proceso arteriosclerótico es rápido y reproducible. Además, si estos ratones se alimentan con una dieta enriquecida en colesterol y grasas, los valores de colesterol plasmático y la aterogénesis aumentan de manera proporcional1.
Las alteraciones del metabolismo de las lipoproteínas son un factor de riesgo para la arteriosclerosis y pueden producirse como resultado de errores en la regulación de las apolipoproteínas. En el cromosoma 11 humano existe una región que agrupa los genes de las apolipoproteínas A-I/C-III/A-IV/A-V, que ha sido muy estudiada por su implicación en el metabolismo lipídico. El último de ellos (APOA5) se ha descubierto recientemente y ha demostrado tener una influencia especialmente relevante en el control de la trigliceridemia. Los animales que sobreexpresan el gen tienen un 60% menos triglicéridos (TG) circulantes, y en los animales que carecen del gen la trigliceridemia aumenta hasta 4 veces2. El gen APOA5 se expresa únicamente en el hígado, donde se postula que reprime la síntesis y/o secreción de las lipoproteínas de muy baja densidad (VLDL)3. Estudios in vitro demuestran que la apo A-V estimula la actividad hidrolítica de la lipoproteína lipasa unida al endotelio4. En los últimos años, varios grupos han descrito (mediante estudios de asociación) que los polimorfismos en el gen APOA5 se asocian a mayores concentraciones de TG en plasma5, aunque su relación con el riesgo cardiovascular es todavía controvertida6-8. Otros estudios describen una asociación entre polimorfismos del gen APOA5 y la dieta y sugieren que su expresión está regulada por la alimentación9,10, aunque este punto no ha sido demostrado.
Nuestro objetivo ha consistido en estudiar el efecto que produce la dieta en la expresión de APOA5 en un modelo experimental de arteriosclerosis y la influencia de los cambios inducidos en APOA5 sobre parámetros lipídicos, la propia lesión arteriosclerótica y las concentraciones circulantes del marcador inflamatorio clave en ratones, el monocyte chemotactic protein -1 (MCP-1).
Material y métodos
Ratones KO-apoE y diseño del estudio
La experimentación con los animales se llevó a cabo en las condiciones que establece la normativa legal vigente (Real Decreto 223/1998) y la tutela de la Comisión de Experimentación científica en animales de nuestra universidad.
Se utilizó 72 ratones macho KO-apoE agrupados en jaulas a 21-23 °C, con un grado de humedad del 50-60%, sometidos a un ciclo de 12 h de luz/oscuridad y con libre acceso tanto al agua como a la comida.
Los ratones se dividieron en 3 grupos de estudio que recibieron dieta convencional de ratón (pobre en grasa y sin colesterol. BK Universal, Barcelona) o dieta rica en grasas saturadas (20% aceite de palma. Unilever, Bilbao) con diferente contenido en colesterol (0 y 0,25% p/p. Sigma Chemical Co., St. Louis, MO, Estados Unidos) y se han sacrificado a las 16, 24 y 32 semanas de edad. Este diseño nos permite estudiar la expresión del gen APOA5 en relación con la dieta y con la edad y, por tanto, observar patrones temporales.
Obtención de muestras
Durante el procedimiento experimental, se obtuvo muestras de sangre mediante punción en la cola y semanalmente se realizó el control del peso de todos los animales. Inmediatamente antes del sacrificio, se administró a los animales 0,1 ml de una mezcla de ketamina/xilacina para anestesiarlos por vía intraperitoneal. Una vez sacrificados, se obtuvo sangre mediante punción ventricular y también se extrajo el hígado y el corazón, que se congelaron inmediatamente a 80 ºC, hasta su posterior utilización.
Determinaciones lipídicas
Se determinaron los valores de colesterol y TG en plasma mediante técnicas estándar de laboratorio. Para caracterizar la composición lipídica de la esteatosis presente en estos animales, se determinó los contenidos de colesterol y TG en homogenizados de tejido hepático después de la extracción de lípidos con isopropilo alcohol-hexano11.
Cuantificación de la lesión ateromatosa
La cuantificación de la lesión ateromatosa se determinó según el método de Tangirala et al12. Después del sacrificio, se congeló inmediatamente el corazón junto con el origen de la aorta; los cortes seriados obtenidos mediante un criostato se han teñido con un colorante con afinidad por los lípidos, el Sudan IV. La imagen microscópica para medir las áreas teñidas con este colorante, correspondientes a las áreas de lesión ateromatosa, se capturó mediante una cámara de vídeo y la extensión de la arteriosclerosis se ha cuantificado con un sistema de análisis de imagen automatizado (AnalySIS Soft Imaging System, Münster, Germany).
Concentración plasmática de MCP-1
La concentración plasmática del marcador inflamatorio MCP-1 se ha determinado utilizando un kit de ELISA (Mouse MCP-1 ELISA kit, Pierce, Rockford, Estados Unidos), siguiendo las instrucciones del fabricante.
Extracción de ARN y PCR a tiempo real
La extracción del ARN total de tejido hepático se realizó mediante el equipo de extracción ABI PRISM 6100 Nucleic Acid PrepStation de Applied Biosystems siguiendo las instrucciones del fabricante. La pureza y la concentración del ARN se han estimado mediante la relación de absorbancia 260 nm/280 nm. Se ha retrotranscrito 1 µg de ARN a ADNc siguiendo las instrucciones del fabricante y utilizando el termociclador PE Biosystems 2400.
La expresión del ARN mensajero (ARNm) de APOA5 se ha cuantificado mediante reacción en cadena de la polimerasa (PCR) a tiempo real utilizando el equipo ABI Prism 5700 Sequence Detector System (Applied Biosystems, Foster City, Estados Unidos), combinado con la utilización de primers y sondas Taqman®. Éstos se obtuvieron de Applied Biosystems como productos Assays-on-Demand validados y prediseñados. El ARN ribosomal (ARNr) A 18S se utilizó como control endógeno y los valores de expresión se calcularon de acuerdo con el método de DDct (ABI Prism 7700 Sequence Detection System. User bulletin n.o 2. Revisión A. Foster city [CA]: Applied Biosystems, 1997).
Análisis estadístico
Los datos se expresan como media ± desviación estándar. La comparación de medias se ha determinado mediante la prueba de ANOVA univariante y para las variables categóricas se utilizó la prueba de la *2.
Las correlaciones entre la expresión de APOA5, la extensión de la lesión en la aorta y la concentración plasmática del parámetro inflamatorio MCP-1 se han determinado mediante regresión lineal.
Resultados
Efecto de la dieta sobre los lípidos y la arteriosclerosis
En la tabla 1 se muestra cómo la administración de dieta rica en grasa saturada (20% aceite de palma), al suplementarla con colesterol, hace aumentar significativamente los valores circulantes de colesterol respecto a la dieta convencional en los ratones KO-apoE (p < 0,0001) y no modifica las concentraciones circulantes de TG. La suplementación con colesterol también produce un aumento significativo de los valores de colesterol y TG en el hígado (p < 0,0001 y p = 0,013, respectivamente).
La superficie de lesión arteriosclerótica medida en la aorta y la presencia de esteatosis en el hígado aumentan significativamente con el contenido en grasa de la dieta (p < 0,0001) (fig. 1). La concentración plasmática de MCP-1 también tiende a aumentar, pero este incremento no es estadísticamente significativo.
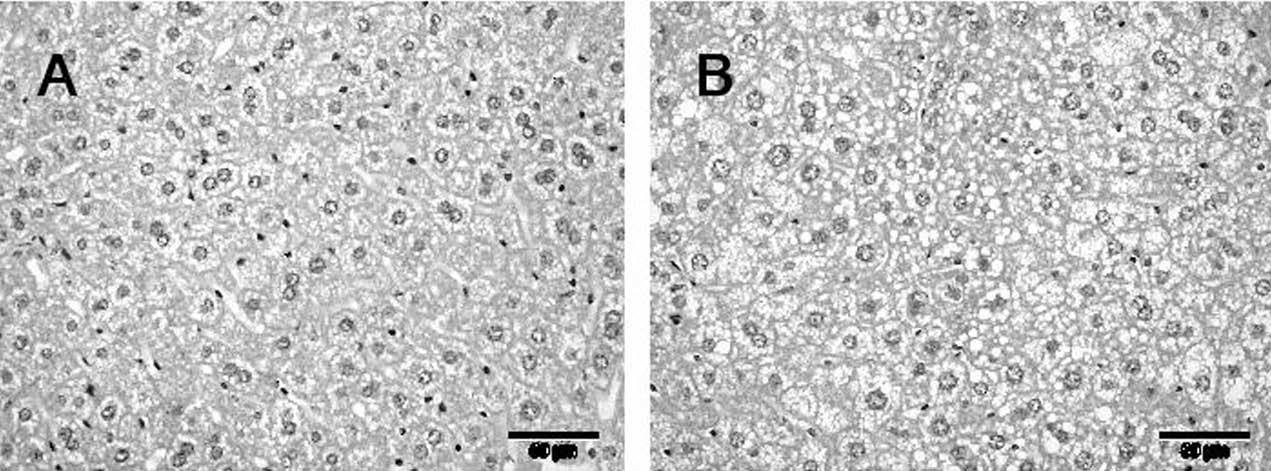
Figura 1. Esteatosis en hígado según el tipo de dieta.
Secciones de tejido hepático de ratón KO-apoE de 32 semanas de edad alimentado con dieta convencional (A) y de ratón alimentado con dieta rica en grasa saturada (B) donde se observa una mayor presencia de microvesículas lipídicas.
La ingesta de grasa saturada disminuye la expresión hepática del gen APOA5
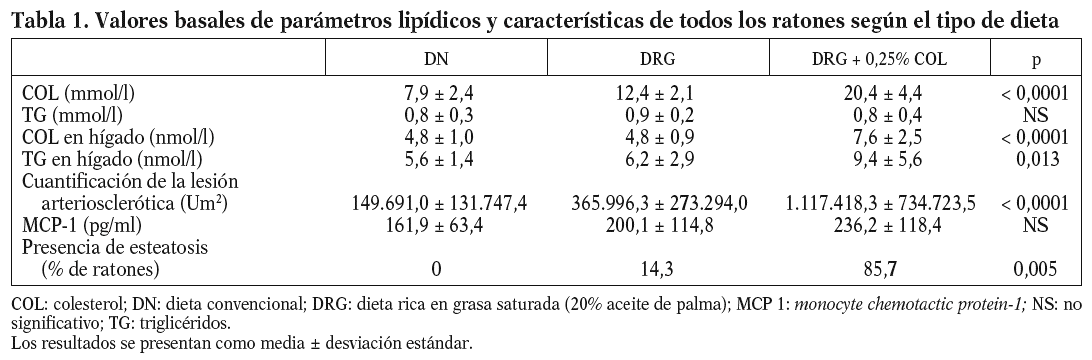
Concretamente, la adición de un 20% de aceite de palma disminuye los valores de ARNm de APOA5 en el hígado en un 75% (p = 0,020) en los ratones de 16 semanas, un 35% (p = NS) en los de 24 semanas y un 45% (p = 0,023) en los de 32 semanas, respecto a la dieta convencional. Sin embargo, la adición de 0,25% de colesterol a la dieta rica en grasa saturada evita esta disminución en los valores de ARNm de APOA5 que no difieren de los de la dieta convencional tanto en los ratones de 16 semanas (p = 0,003) como en los de 32 (p = 0,039) (fig. 2).
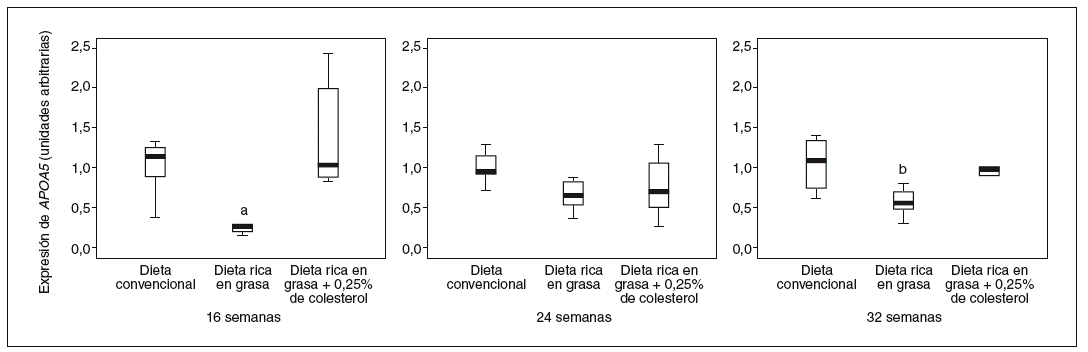
Figura 2. Distribución de los valores de expresión hepática de APOA5 en relación con el tipo de dieta que recibían los ratones agrupados por edad. La expresión hepática de APOA5 en los ratones alimentados con dieta rica en grasa saturada está significativamente disminuida respecto de la que presentan los ratones alimentados con dieta convencional o dieta rica en grasa saturada suplementada con colesterol en los ratones de 16 (p = 0,020)a, 24 (p = NS) y 32 (p = 0 ,023)b semanas de edad, respectivamente.
Efecto de la edad sobre los lípidos y la arteriosclerosis
La tabla 2 muestra las diferencias entre los parámetros lipídicos determinados en los ratones alimentados con todas las dietas, según la edad a la que fueron sacrificados.

Vemos que con la edad los valores circulantes de colesterol no presentan ningún cambio y que los valores de TG circulantes disminuyen significativamente a las 32 semanas (p = 0,016). Los valores de colesterol en el hígado tienden a disminuir con la edad y en cuanto a los TG vemos que disminuyen significativamente (p = 0,003). El área de la lesión arteriosclerótica aumenta, tal y como esperábamos, así como el grado de inflamación determinado con la concentración de MCP-1 en plasma (p < 0,0001 y p < 0,0001, respectivamente). Con la edad también aumenta el número de ratones que presentan esteatosis (p = 0,037).
La expresión hepática del gen APOA5 aumenta con la edad
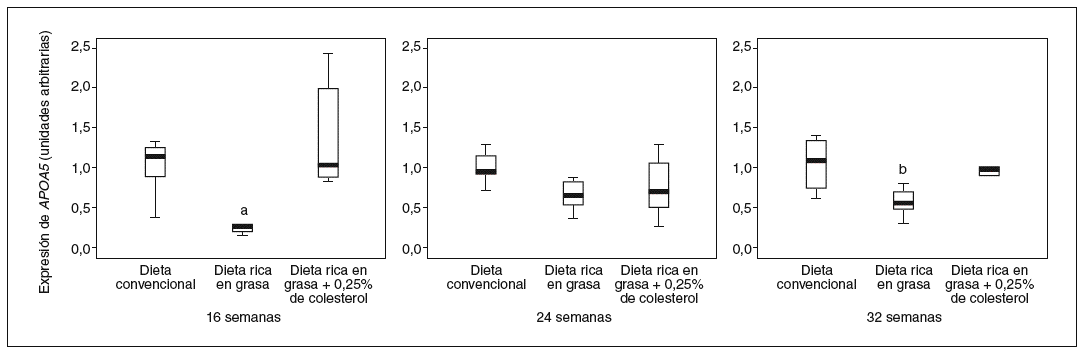
Los valores hepáticos de ARNm de APOA5 se correlacionan positivamente con la edad (R2 = 0,405; p < 0,0001) y negativamente con los valores de colesterol y TG en el hígado (R2 = 0,082; p = 0,047; y R2 = 0,087; p = 0,040, respectivamente). Los ratones de 24 y 32 semanas de edad presentan unos valores de media del 89,1 y el 282,8% superiores respecto a la expresión que presentan los de 16 semanas (fig. 3). Este efecto es independiente de la dieta ya que también se observa si separamos los ratones en los diferentes grupos de dieta.
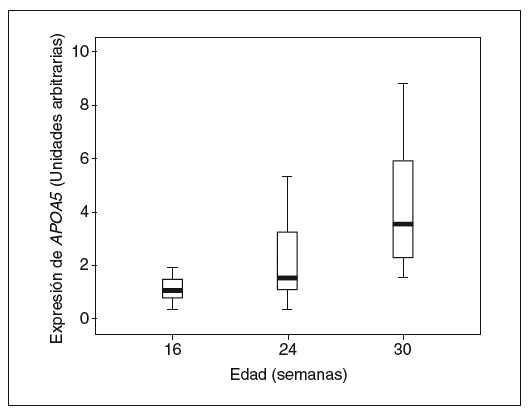
Figura 3. Distribución de los valores de expresión hepática de APOA5 en relación con la edad a la que fueron sacrificados los ratones. Los ratones de 32 semanas de edad presentaban mayores valores de expresión de APOA5 en hígado respecto de los ratones de 16 semanas (p < 0,001).
Arteriosclerosis
Para intentar comprender el alcance de los cambios en APOA5 inducidos por las distintas dietas, analizamos las correlaciones entre los valores de ARNm de APOA5 y de lípidos, la lesión ateromatosa y el grado de inflamación en estos ratones.
La expresión hepática de APOA5 no se correlaciona significativamente con ningún parámetro lipídico determinado en los ratones.
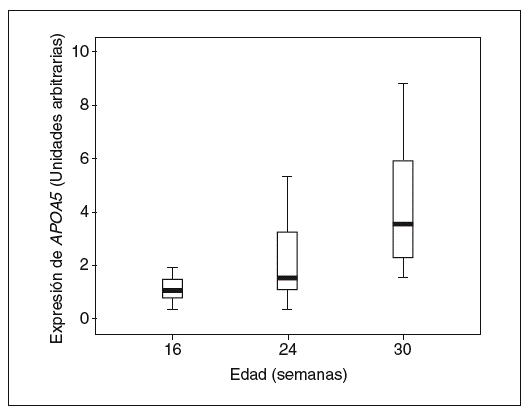
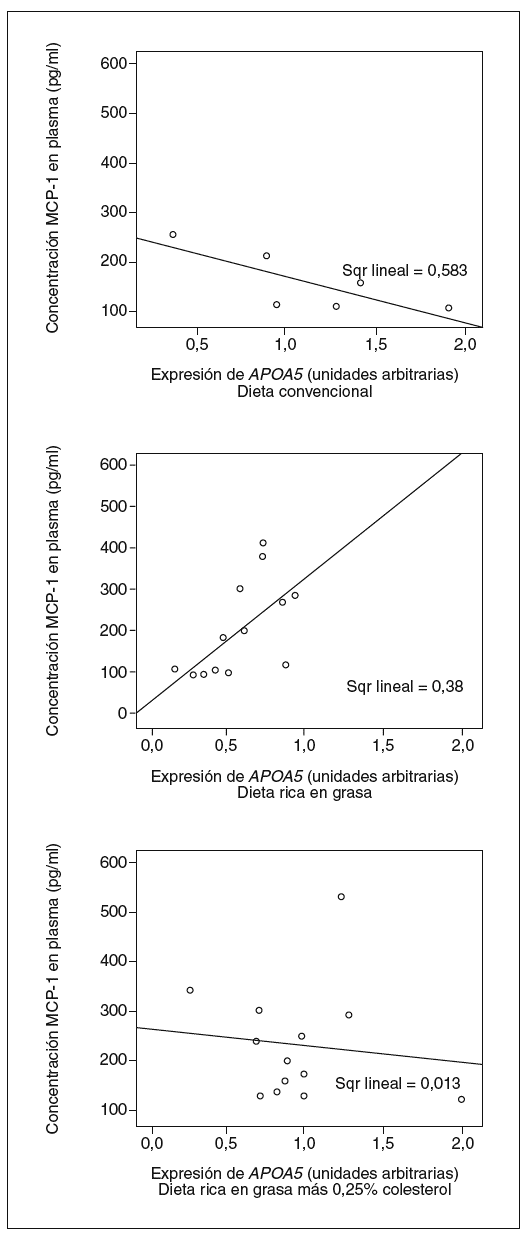
Tal y como muestran las figuras 4 y 5, observamos que la expresión de APOA5 tiende a correlacionarse positivamente con la superficie de lesión en la arteria aorta de los ratones alimentados con dieta rica en grasa saturada, y negativamente en los ratones alimentados con dieta convencional o dieta rica en grasa saturada suplementada con colesterol. En el caso de la concentración plasmática de MCP-1, también observamos el mismo comportamiento de forma positiva y significativa en los ratones alimentados con dieta rica en grasa saturada y correlacionando negativamente en los ratones alimentados con los otros 2 tipos de dietas.
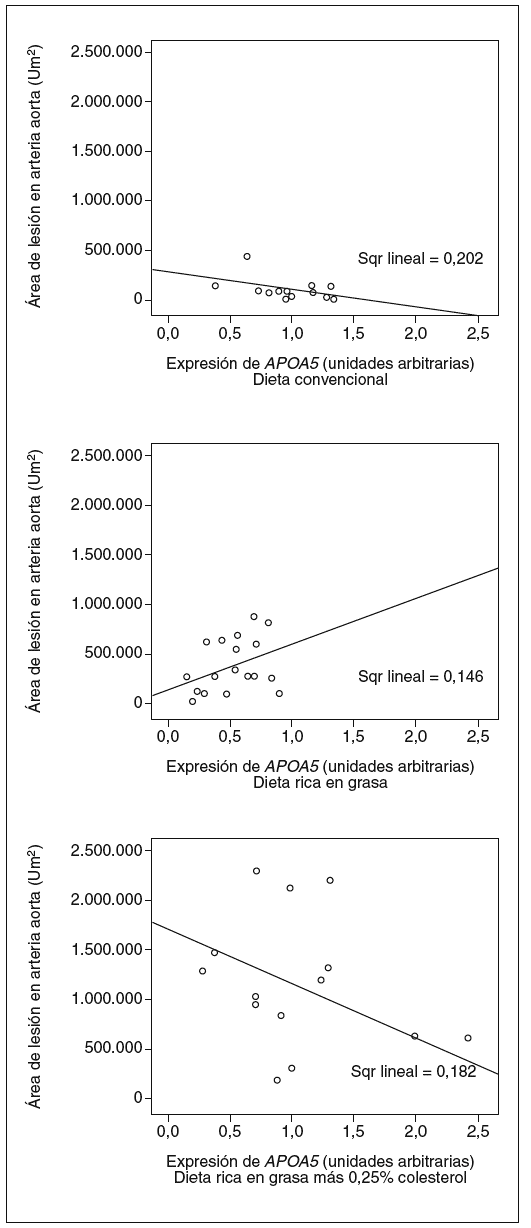
Figura 4. Correlación entre la expresión hepática de APOA5 y la cantidad de lesión en la arteria aorta según el tipo de dieta con que estaban alimentados los ratones. Los ratones alimentados con dieta convencional presentan una correlación negativa con la superficie de lesión en la arteria aorta (R2 = 0,202; p = NS), al igual que los alimentados con dieta rica en grasa más 0,25% colesterol (R2 = 0,182; p = NS). En cambio, los ratones alimentados con dieta rica en grasa saturada tendían a presentar una correlación positiva con el área de la lesión (R2 = 0,146; p = NS).
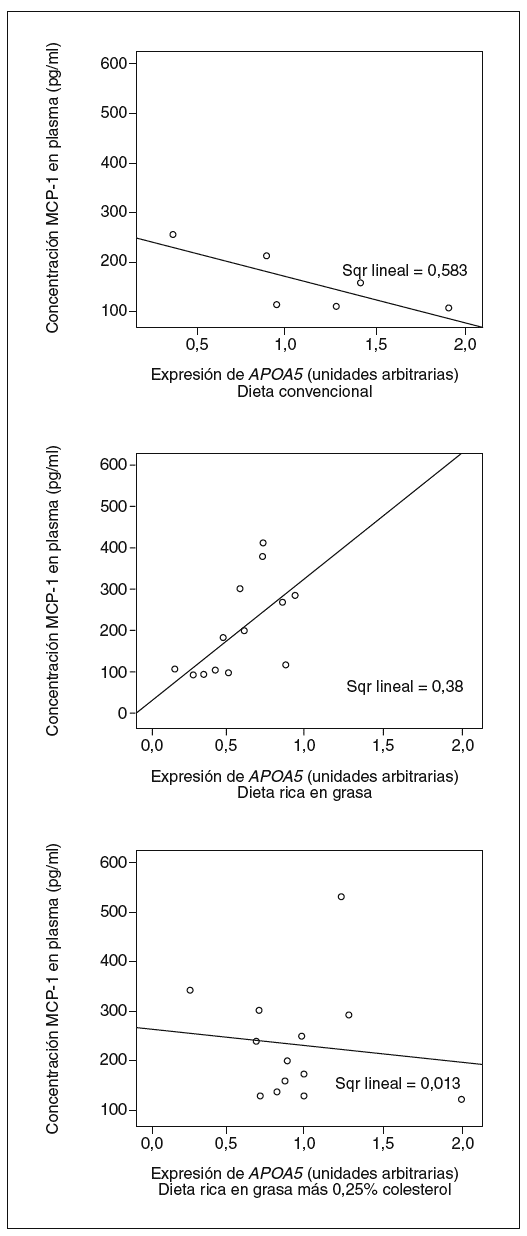
Figura 5. Correlación entre la expresión hepática de APOA5 y la concentración plasmática de monocyte chemotactic protein 1 (MCP-1) según el tipo de dieta con que estaban alimentados los ratones. Los ratones alimentados con dieta convencional tendían a presentar una correlación negativa con la concentración de MCP-1 en plasma (R2 = 0,583; p = NS), al igual que los alimentados con dieta rica en grasa más 0,25% colesterol (R2 = 0,013; p = NS). En cambio, los ratones alimentados con dieta rica en grasa saturada presentaban una correlación positiva con el área de la lesión (R2 = 0,380; p = 0,025).
Discusión
Nuestros resultados demuestran que la grasa saturada disminuye cerca de un 50% la expresión hepática de APOA5 en el ratón KO-apoE. Además, aumenta con la edad en todas las dietas administradas.
Dieta
La expresión hepática del gen APOA5 disminuye significativamente en todos los ratones de diferentes edades alimentados con dieta rica en ácidos grasos saturados; sin embargo, al suplementar esta dieta con 0,25% de colesterol tal efecto desaparece.
Los modelos animales indican que la sobreexpresión de APOA5 ayuda a controlar los valores plasmáticos de TG13. Por lo tanto, la disminución de hasta un 75% en animales jóvenes de la expresión hepática de APOA5 supondría un efecto adverso en los TG. Ello estaría de acuerdo con los estudios que muestran que una dieta rica en grasa saturada aumenta los valores de TG y que el déficit de apo A-V en humanos está asociado a hipertrigliceridemia grave14,15. Sorprendentemente, no hemos observado que la disminución de APOA5 afecte a los valores de TG. Creemos que ello puede ser una característica específica de los ratones KO-apoE en los que sabemos que la vía secretora de TG en los hepatocitos está alterada16, lo cual podría restar influencia a APOA5 en la síntesis de VLDL.
El mecanismo mediante el cual los ácidos grasos libres y el colesterol actuarían en APOA5 no está claro; sin embargo, sabemos que los ácidos grasos regulan la expresión de genes implicados en el metabolismo energético y en este proceso de regulación también están implicados varios tipos de receptores nucleares encargados del reconocimiento de secuencias en la zona reguladora del gen. En el caso de APOA5, sabemos que está regulado, entre otros, por receptor de la activación de peroxisomas * (PPAR*)17, que tiene como ligandos naturales los ácidos grasos, y nuestros resultados describen por primera vez un papel regulador de los ácidos grasos saturados en la expresión de APOA5.
El efecto de la suplementación de colesterol en la expresión hepática de APOA5 puede explicarse teniendo en cuenta que su expresión está estimulada por el receptor nuclear ROR*18,19, cuya actividad transcripcional está regulada por el colesterol. Está descrito que las condiciones que afectan al contenido celular de colesterol pueden modular la actividad de ROR* in vitro20, por tanto, es de esperar que el tipo de dieta que administramos en estos ratones, que aumenta los valores de colesterol, hace que ROR* sea constitutivamente activo y, por tanto, estimule la expresión de APOA5, por lo que se pierde el efecto producido por la dieta rica en grasa saturada.
Edad
El ratón KO-apoE alimentado con dieta aterogénica experimentan un deterioro progresivo del hígado (esteatosis e inflamación). La expresión he pática del gen APOA5 está significativamente aumentada con la edad y este efecto no depende del tipo de dieta recibida. Sin embargo, debido al deterioro funcional del hígado de estos animales sometidos a dietas aterogénicas, creemos que este aumento puede deberse a este hecho más que a la edad per se. Además, los cambios en la expresión de APOA5 debidos a la edad se correlacionan de forma negativa y significativa con los valores de colesterol y TG en el hígado. Ello es congruente con el hecho de que APOA5 es uno de los genes más sobreexpresados durante el proceso de regeneración hepática y que ya ha sido considerado como proteína de fase aguda21,22.
Arteriosclerosis
Desde que se descubrió en 2001 el gen APOA5 se ha asociado a mayores valores de TG circulantes en varios tipos de poblaciones como diabéticos e hiperlipémicos; nuestro grupo ha descrito asociación en pacientes con HLFC y en pacientes infectados por el virus de la inmunodeficiencia humana tratados con inhibidores de la proteasa23-25. También se ha asociado a mayor riesgo de enfermedad cardiovascular y se ha identificado como proteína de fase aguda, un efecto independiente de los valores plasmáticos de TG.
Como ya se ha comentado, el descenso en la expresión hepática de APOA5 debido a la ingesta de dieta rica en grasa saturada no se correlacionaba con los valores de lípidos, posiblemente debido a la alteración en la vía secretora hepática de TG en los ratones KO-apoE16. Por ello nos interesó saber si este efecto se correlacionaba con otros parámetros implicados con el proceso arteriosclerótico determinados en estos ratones.
La expresión hepática de APOA5 se correlaciona con parámetros inflamatorios como la concentración en plasma de MCP-1 y con la superficie de lesión arteriosclerótica. Estas correlaciones varían según el tipo de dieta con que alimentamos a los ratones, un hecho que refleja la importancia del efecto modulador de la dieta en la enfermedad cardiovascular y sus factores de riesgo. Según el tipo de dieta, observamos que los valores de expresión hepática de APOA5 presentan un patrón de expresión inverso. Con la dieta convencional de ratón, los valores de expresión de APOA5 se correlacionan negativamente con la cantidad de lesión arteriosclerótica y con el grado de inflamación presente en el ratón, lo que sugiere cierta asociación con el riesgo cardiovascular. Con la dieta rica en grasa saturada vemos que se correlaciona positivamente, tanto con el grado de arteriosclerosis como con el grado de inflamación de los ratones, y probablemente actúa como proteína de fase aguda en condiciones de estrés del organismo.
En conjunto, los datos son congruentes con una doble función del gen de APOA5. Como regulador del metabolismo de los lípidos, en relación con la síntesis y secreción de lipoproteínas y el control de las reservas hepáticas de grasa, y como proteína de fase aguda con unos valores más elevados de actividad transcripcional, asociados a parámetros inflamatorios.
En resumen, la grasa saturada de la dieta disminuye cerca de un 50% la expresión hepática de APOA5 en el ratón KO-apoE, y por tanto, puede representar una forma de modificar la expresión de APOA5 en el hígado.