









An analysis of wastewater samples collected from different industrial regions of Egypt demonstrated dangerously high levels of nickel (0.27–31.50mgL−1), chromium (1.50–7.41mgL−1) and zinc (1.91–9.74mgL−1) in the effluents. Alarmingly, these heavy metals are among the most toxic knownones to humans and wildlife. Sixty-nine Actinomycete isolates derived from contaminated sites were evaluated under single, binary, and ternary systems for their biosorption capacity for Ni2+, Cr6+ and Zn2+ from aqueous solutions. The results of the study identified isolates MORSY1948 and MORSY2014 as the most active biosorbents. Phenotypic and chemotypic characterization along with molecular phylogenetic evidence confirmed that the two strains are members of the Nocardiopsis and Nocardia genera, respectively. The results also proved that for both the strains, heavy metal reduction was more efficient with dead rather than live biomass. The affinity of the dead biomass of MORSY1948 strain for Ni2+, Cr6+ and Zn2+ under the optimized pH conditions of 7, 8 and 7, respectively at 40°C temperature with 0.3% biosorbent dosage was found to be as follows: Ni2+ (87.90%)>Zn2+ (84.15%)>Cr6+ (63.75%). However, the dead biomass of MORSY2014 strain under conditions of pH 8 and 50°C temperature with 0.3% biosorbent dose exhibited the highest affinity which was as follows: Cr6+ (95.22%)>Ni2+ (93.53%)>Zn2+ (90.37%). All heavy metals under study were found to be removed from aqueous solutions in entirety when the sorbent dosage was increased to 0.4%.
The most significant sources of heavy metal contamination are human activities and industries such as electroplating, electroforming, painting, petrochemicals, chemical manufacturing, pigments, rechargeable batteries, electronics and computer equipment, metals (coatings, plating and finishing operations), steel, manufacturing detergents, and coins. The discharge of untreated metal-containing effluent into the natural environment in quantities that exceed prescribed limits is becoming an issue of great concern and is causing consternation to environmentalists as well as government agencies.1,2 The examples of such metals that are known to be significantly toxic to humans as well as the ecological environment include chromium (Cr6+), nickel (Ni2+), zinc (Zn2+), copper (Cu2+), lead (Pb2+), cadmium (Cd2+), and mercury (Hg2+). Technologies for removing heavy metals, including methodologies such as chemical precipitation, ion-exchange, reverse osmosis, electro-dialysis, and ultra-filtration, are routinely being used for treating industrial wastewater but because they suffer from disadvantages such as being extremely expensive while being inefficient at metal removal and because they result in generation of toxic compounds, they are now being seen as both uneconomical and unfavorable.3 There is, therefore, a pressing need for the development of highly selective yet cheap and efficient alternatives that can mitigate heavy metal concentrations in wastewater to environmentally accepted levels. One such promising methodology being developed is the bioremediation of heavy metals from aqueous solutions using certain specific types of metabolically active (live cells) or inactive (dead cells) microbial biomasses.4 Studies have indicated that Actinomycetes as biosorbents are both efficient as well as economical in treating effluents and removing toxic metals from waste water; this property is attributed to the presence of a large number of functional groups on their cell walls and their filamentous morphology.4–7
The present study is aimed assessing the ability of Actinomycetes to remove toxic heavy metals such as Ni2+, Cr6+ and Zn2+, from aqueous solutions. It is also an objective of this study to improve the adsorption capacity of selected biosorbent strains by optimizing the removal parameters (pH, temperature, biomass nature, dead or alive, biomass dosage, heavy metal concentrations versus different contact time). We also aim to determine the desorption and recovery efficiency of these metals from the biosorbent biomasses and to determine the metal toxicity and regeneration ability of Actinomycetes following exposure to a wide range of heavy metals that might be present in real industrial wastewater.
Materials and methodsChemicals and factory effluentsDeionized water was used for the preparation of standard heavy metal solutions (concentration: 2gL−1). Freshly diluted solutions of varying concentrations (mgL−1: 50, 100, 200, 300, 400, and 500) that were used in all experiments were prepared by serially diluting with deionized water. Binary and ternary metal solutions were prepared using 100mgL−1 for each one of the metals under study and mixing the same in equal proportions. The pH of the metal ion solutions was adjusted to the desired values using either concentrated HNO3 (65%) or 1M NaOH.8
Effluents belonging to a variety of industries were collected from ten different industrial regions of Egypt (10th of Ramadan, Gesr El Suez, Badr city, 6th of October, Shubra El-Kheima, Sadat city, Borg El-Arab, Abu-Rawash, free zone-Nasr city and El-Amerya, Egypt). The collected wastewater samples were filtered and placed in sterile 250mL conical flasks containing 2.5mL nitric acid (conc.). These flasks were kept at 4°C until analyzed for their Ni2+, Cr6+ and Zn2+ contents. This assessment, conducted within 24h of collection, was executed by Atomic Absorption Spectrophotometer (AAS, Model-M series Thermo-Scientific, NIOSH) as previously reported by El-Gendy et al.9
Isolation and identification of Actinomycetes biosorbentsActinomycetes biosorbents were isolated from environments that are naturally rich in heavy metals such as polluted soils (10th of Ramadan and 6th of October, Egypt) and water drainage areas such as the Nile Delta in Lower Egypt. For soil samples, about 100g of air dried soil was collected in plastic bags; polluted water samples were collected in screw cap test tubes and stored at 4°C to stop any biological activity until processing (24h). For isolation of Actinomycetes, polluted water drainage samples were filtered and inoculated into Actinomycetes isolation medium using the serial dilution technique. Soil samples were prepared by heat treatment at 60°C for 45min followed by overnight extraction in 600mL distilled water by shaking. The samples were then centrifuged for 10min at 4000rpm; and 100μL aliquots of 10−3 to 10−5 dilutions of the supernatant were serially diluted and plated on soil extract agar.10 The plates were incubated for 7 days at 28°C and Actinomycetes colonies were identified visually by their tough leathery appearance. The presence of branched vegetative mycelia with aerial mycelia and spore formation were microscopically analyzed and isolates fulfilling all criteria were transferred periodically to tryptic soy agar medium. The 69 Actinomycete isolates obtained were analyzed for their biosorption efficiency and the most active biosorbent isolates, MORSY1948 and MORSY2014 were selected and identified as previously described.11–25
Molecular identificationGenomic DNA extraction, PCR amplification of the 16S rDNA, the purification of PCR products, gel electrophoreses and 16S rDNA sequencing were done as described previously.26,27 The 16S rDNA sequences of the most promising biosorbent isolates, MORSY1948 and MORSY2014, were aligned with the published representative sequences of Actinomycetes obtained from the NCBI GenBank and DSMZ database for 16S rDNA sequences. The tree topologies were evaluated by maximum-likelihood and bootstrap analysis based on 1000 replications with MEGA6, and phylogenetic trees were inferred using the neighbor-joining method.28,29 The complete 16S rDNA sequences of the MORSY1948 and MORSY2014 strains were deposited in the GenBank database under accession numbers: KP979750 and KP979751, respectively.
Preparation of live and dead biomassesThe live and dead biomasses of the Actinomycete isolates were used individually as natural biosorbents to test the bioremoval of Ni2+, Cr6+ and Zn2+ from aqueous solutions. Ten-day old culture spores (106CFUmL−1) of each Actinomycete isolate were transferred individually into 500mL Erlenmeyer flasks containing 100mL broth medium [(gL−1): peptone, 4; yeast extract, 2; glucose, 10] and incubated at 30°C, 150rpm on a rotary shaker for 7 days. Thereafter, the resultant biomass of each Actinomycete isolate was pelletized by filtration through filter papers (Whatman No. 1), washed five times with 0.1M NaCl followed by sterile distilled water in order to remove non biomass particles. Dead biomass experiments were performed as described previously by Vijayaraghavan and Yun.30 The inactive cells were washed with 0.1M NaCl following which they were transferred to pre weighted aluminum foil caps and dried in an oven at 60°C until constant weight was obtained. To assess complete death of the dried cells, the samples were inoculated into Petri dishes containing the medium described previously; absence of any growth was presumed to be indicative of positive results. Live biomass was obtained by air drying the cells followed by pulverizing them to a fine powder using a porcelain mortar. The concentrations of both live and dead biomasses were calculated subsequent to which they were stored at 4°C till required.31,32
Evaluation of metal biosorption capacity of Actinomycete isolatesUnless stated otherwise, the biosorption tests were conducted using quick-fit flasks containing 3gL−1 dead biomass of the individual Actinomycete sorbent under study (biosorbent dosage) in 100mL aliquots of metal solution containing 100mgL−1 of one of the metal ions of interest. Flasks were kept on rotary shakers (150rpm) at 30°C and pH 6.0 for 3h. Subsequently, the samples were centrifuged at 10,000rpm for 15min. The supernatants were analyzed for residual heavy metals using Atomic Absorption Spectrophotometer. Presence of Ni2+, Cr6+ and Zn2+ was determined by using different lamps specific for each metal at specific wavelengths. Metal solutions without biomass addition served as control. Experiments were conducted in duplicate and average values were computed. The following equation was used to compute the in-solution metal biosorption efficiency (R) for each metal ion by each isolate; and the results were expressed in percentage terms: Percent Biosorption (R)=(Ci−Cf)/Ci×100, where Ci corresponds to the initial metal ion concentration of the aqueous solution and Cf corresponds to the residual concentration. In addition, metal biosorption calculations for each metal under binary and ternary conditions and in different combinations or in real wastewater conditions were calculated by the following equations: R1 (%)=(C1i−C1f)/C1i×100, R2 (%)=(C2i−C2f)/C2i×100 and R3 (%)=(C3i−C3f)/C3i×100, where R1, R2 and R3 are the biosorption efficiencies of the first, second and third metal respectively (%); C1i, C2i and C3i are initial concentration of first, second and the third metal, respectively (mgL−1) and C1f, C2f and C3f are final, post-biosorption, concentrations of the first, second and third metal respectively (mgL−1).33
Factors affecting the efficiency of the Ni2+, Cr6+ and Zn2+ bioremoval process by MORSY1948 and MORSY2014 isolatesFor analyzing the impact of pH, biosorption by MORSY1948 and MORSY2014 was carried out with varying pH values (2.0, 4.0, 6.0, 7.0, 8.0, 9.0, 10 and 11) under conditions in which 3gL−1 biomass was dispersed in 100mL of a solution containing 100mgL−1 of individual metal of interest. The experiment was kept at continuous shaking (150rpm) for 3h at 30°C following which the aqueous solutions were centrifuged and each supernatant was analyzed for residual metal concentration.
For analyzing the effect of temperature, experiments were conducted at different temperature points (25, 30, 35, 40, 45, 50, 55, 60 and 65°C) under optimum pH following which the samples were analyzed for residual metal concentration as described above.
Different weights of biomass, ranging from 0.05% to 0.5%, were dispersed in each metal solution under optimized parameters to determine conditions for maximum metal ion biosorption. Flasks were left for equilibration and then the solutions were centrifuged (10, 000rpm; 15min) and the final concentration of each metal and its biosorption efficiency (%) were determined using the procedures described earlier.
The effect of initial metal concentration (50, 100, 200, 300, 400 and 500mgL−1 of Ni2+, Cr6+ and Zn2+, separately) was studied by analyzing biosorption under conditions wherein all the parameters (pH, temperature, and biosorbent dosage) were optimum for each strain. Flasks were allowed to attain equilibrium on the rotary shaker and samples were collected at regular time intervals (10, 30, 60, 120, 180, 240, 300, 360, 420 and 1440min) in order to determine bioremoval efficiency (%).
Desorption experimentsTo evaluate the desorption efficiency for each heavy metal, Ni2+, Cr6+ and Zn2+ loaded biomasses were dried at 60°C for 24h after attaining equilibrium of sorption at optimum conditions. The exposure of the dried biomass to 1M H2SO4 for 4h in each cycle allowed the heavy metal to be released. Thereafter, the desorbed metal was analyzed and desorption efficiency was calculated as described by Chu et al.34
Determination of metal toxicityIn order to determine the toxic effects of the various metal ions (Ni2+, Cr6+, and Zn2+) on the growth of biosorbent strains MORSY1948 and MORSY2014, and also to deduce their tolerance toward other heavy metals that might be found present in contaminated sites and industrial wastewaters, spores suspension (107CFUmL−1) of each isolate was individually inoculated onto variety of cultivation media. The culture media used were as follows: (a) starch casein medium (gL−1: starch 10, casein powder 1), (b) St 1 medium (gL−1: peptone 15, yeast extract 3, NaCl 6, glucose 15), (c) tryptic soy broth medium (TSB, gL−1: tryptone 17, soytone 3, dextrose 2.5, NaCl 5, K2HPO4 2), and (d) the modified Kuster's medium (gL−1: glycerol 10, casein 0.3, KNO3 2, K2HPO4 2, NaCl 2, MgSO4 0.05, CaCO3 0.02). For the purpose of the experiment the different media were supplemented with individual sterile filtered metal ions, concentrations ranging from 50 to 1500μgmL−1; some of the heavy metals included Ni2+, Zn2+, Cr6+, Fe3+, Cu2+, Cd2+, Pb2+, Co2+, Hg2+, Mn2+ and Ar2+. Heavy metal concentrations that are required for 50% inhibition (IC50) and the minimum inhibitory concentration (MIC) were determined. Controls were prepared by inoculating the same media but without any metal supplementation. Cultures were incubated at 28°C till growth yield in control flasks was maximal and no further increase in their levels was observed; at this point biomass was quantified for all cultures. The regeneration ability of both MORSY1948 and MORSY2014 strains were tested by subculturing them from the heavy metals treated cultures into non-heavy metals media. For analysis purposes, the value for biomass production in metal treated cultures was expressed as a percentage of that obtained in untreated control cultures; the later were considered as 100% as per the method described by Deepika and Kannabiran35 and El-Gendy et al.9
Results and discussionAnalysis of industrial wastewater samples from different industrial regions of EgyptThe analysis of effluents taken from different factories representing different industrial regions of Egypt (10th of Ramadan, Gesr El Suez, Badr city, 6th of October, Shubra El-Kheima, Sadat city, Borg El-Arab, Abu-Rawash, freezone – Nasr city and El-Amerya) showed that Ni2+ concentration in these effluents ranged between 0.27–31.50mgL−1 and that the highest Ni2+ concentration values (18.41–31.50mgL−1) were detected in the effluents in regions wherein electromechanical, electrical, and steel industries were located; the lowest concentrations were detected in regions where food industries were based (0.27 and 0.85mgL−1, Table 1). It was observed that Cr6+ concentration in these effluents ranged from 1.50 to7.41mgL−1 but that of Zn2+ ranged between 1.91 and 9.74mgL−1. Whereas the highest amounts of Cr6+ were detected in the industrial wastewater of electrical and painting industries (7.41 and 7.25mgL−1, respectively), the highest amounts of Zn2+ were detected in the effluents of painting and chemical & medical industries (9.74 and 6.51mgL−1, respectively; Table 1). As per the threshold limit (TLV), governing the discharge of industrial effluents into underground reservoirs (Nile branches or canals), sewage and coastal environment, the content of Ni2+, Cr6+ and Zn2+ must not exceed (0.1, 0.05 and 1mgL−1, Law 48/82), (0.1, 0.05 and 2mgL−1, Law 4/94) and (1.0, 0.05 and 2mgL−1, Law 44/2000). As a result of this treatment of industrial wastewaters in order to decrease the environmental load is imperative for protecting both the environment as well as public health. Previous studies have also reported dangerously high concentrations of toxic metals ions such as mercury, chromium, cadmium, zinc, copper, lead, and nickel in industrial wastewaters and effluents.4,9,36,37
Analysis of industrial wastewater collected from different industrial regions of Egypt.
| Industry | Region | Metal analysis (mgL−1) | ||
|---|---|---|---|---|
| Ni2+ | Cr6+ | Zn2+ | ||
| Steel Wool | 10th of Ramadan | 25.16 | 3.1 | 5.84 |
| High voltage | 10th of Ramadan | 31.50 | 2.0 | 6.01 |
| Electric | 10th of Ramadan | 18.22 | 3.15 | 3.15 |
| Painting | 10th of Ramadan | 2.09 | 7.25 | 9.74 |
| Food | 10th of Ramadan | 0.85 | 1.80 | 1.91 |
| Steel | Gesr El Suez | 20.25 | 4.2 | 6.05 |
| Precise industries | Gesr El Suez | 11.38 | 3.0 | 4.00 |
| Printing | Gesr El Suez | 6.82 | 2.90 | 4.02 |
| Chemical and medical | Badr City | 4.59 | 4.13 | 6.51 |
| Electromechanical engineering | 6th of October | 29.16 | 5.06 | 3.00 |
| Electric | Shubra El-Kheima | 18.41 | 7.41 | 4.83 |
| Electric | Sadat City | 24.36 | 3.98 | 2.77 |
| Electric | Borg El-Arab | 17.91 | 4.25 | 5.47 |
| Chloride | Abu Rawash | 8.15 | 4.99 | 5.00 |
| Medical supplies | free zone, Nasr city | 2.90 | 2.80 | 6.10 |
| Food | El-Amerya | 0.27 | 1.50 | 2.00 |
| Threshold limit values of discharge heavy metals into: | ||||
| Underground reservoir, Nile and canals (Law 48/82) | 0.1 | 0.05 | 1 | |
| Sewage system (Law 44/2000) | 1.0 | 0.05 | 2 | |
| Coastal environment (Law 4/94) | 0.1 | 0.05 | 2 | |
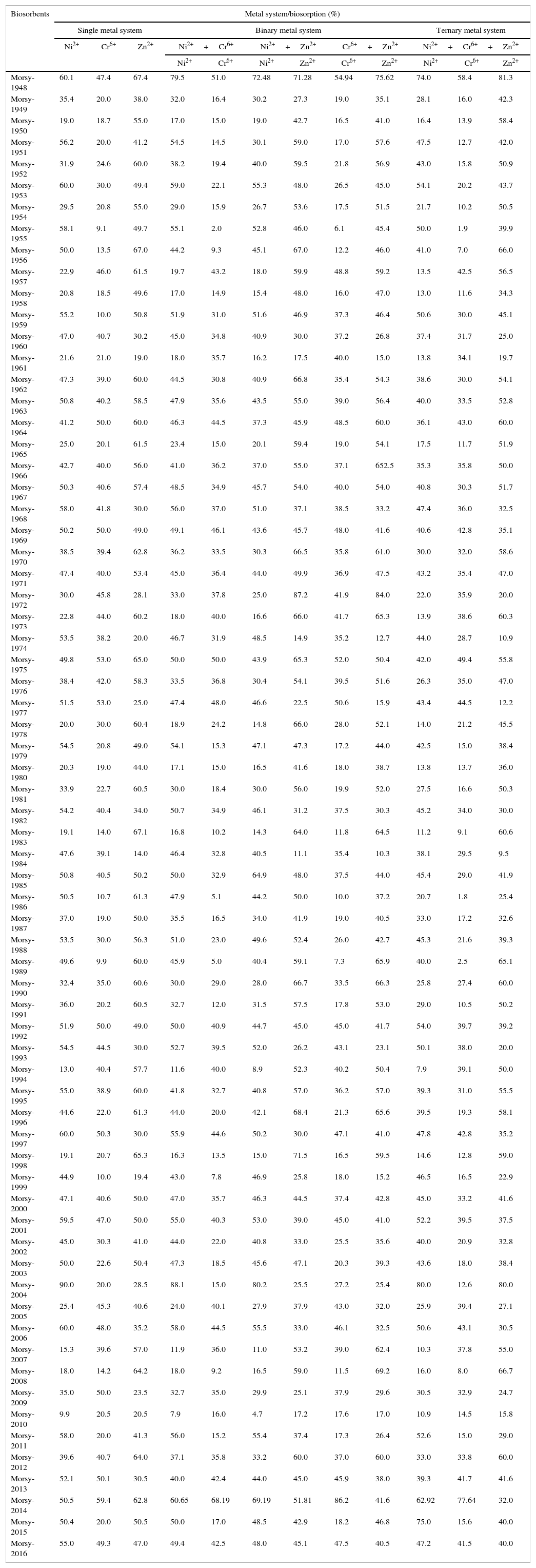
All Actinomycete isolates (69) obtained from polluted sites were found to remove appreciable amounts of the heavy metals under study (Ni2+; Cr6+ and Zn2+) as is clearly evident from the biosorption values (%) of these metals (Table 2). Among all the isolates that were tested, it was found that the biomass of isolate MORSY1948 supported the highest removal of Zn2+ (67.4%) followed by Ni2+ (60.1%) along with significant removal of Cr6+ (47.4%). But it was seen that the binding sites in the biomass of MORSY2014 exhibited the highest removal efficiency for Cr6+ (59.4%) followed by Zn2+ (62.8%) and Ni2+ (50.5%). This behavior of the MORSY2014 strain can be explained by its superior ability to sequester substantial amounts of the heavy metals from aqueous solution when compared to the other isolates. Both the strains, MORSY1948 and MORSY2014, were selected for further studies. It is well established that adapted microbial populations are prone to exhibit higher resistance to heavy metals as compared to populations of non-contaminated sites.9,36 Reports published by previous studies have supported the role of Actinomycetes in the removal of toxic metals from the contaminated soil; examples include: Streptomyces coelicolor (Cu2+), S. pimprina (Cd2+), S. rimosus (Cd2+, Pb2+, Cu2+, Zn2+, Fe3+ and Cr6+), Streptoverticillium cinnamoneum (Pb2+), S. rimosus (Fe3+), S. rimosus (Ni2+), Nocardia erythropolis IAM 1399 (Th4+ and U6+), S. rimosus and Streptoverticillium cinnamoneum (Zn2+), Actinomycete strains (Cr6+), Streptomyces sp. 19H (Au2+), Arthrobacter species (Cr3+ and Cr6+).1,6,36,38,39 It is hypothesized that the superior metal adsorbing capacity of Actinomycetes might be due to the relatively high phosphorus content of their cell wall, as it is known that the major metal binding site is the teichoic acid moiety.5,31
Comparative representation of biosorption efficiency (%) of various Ni2+, Cr6+, Zn2+ heavy metals under different metal systems by Actinomycete biosorbent isolates (biosorption conditions are 100mgL−1 of such heavy metal, 0.3% of biosorbent, pH 6.0 at 30°C and 150rpm for 3h).
| Biosorbents | Metal system/biosorption (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Single metal system | Binary metal system | Ternary metal system | ||||||||||
| Ni2+ | Cr6+ | Zn2+ | Ni2++Cr6+ | Ni2++Zn2+ | Cr6++Zn2+ | Ni2++Cr6++Zn2+ | ||||||
| Ni2+ | Cr6+ | Ni2+ | Zn2+ | Cr6+ | Zn2+ | Ni2+ | Cr6+ | Zn2+ | ||||
| Morsy-1948 | 60.1 | 47.4 | 67.4 | 79.5 | 51.0 | 72.48 | 71.28 | 54.94 | 75.62 | 74.0 | 58.4 | 81.3 |
| Morsy-1949 | 35.4 | 20.0 | 38.0 | 32.0 | 16.4 | 30.2 | 27.3 | 19.0 | 35.1 | 28.1 | 16.0 | 42.3 |
| Morsy-1950 | 19.0 | 18.7 | 55.0 | 17.0 | 15.0 | 19.0 | 42.7 | 16.5 | 41.0 | 16.4 | 13.9 | 58.4 |
| Morsy-1951 | 56.2 | 20.0 | 41.2 | 54.5 | 14.5 | 30.1 | 59.0 | 17.0 | 57.6 | 47.5 | 12.7 | 42.0 |
| Morsy-1952 | 31.9 | 24.6 | 60.0 | 38.2 | 19.4 | 40.0 | 59.5 | 21.8 | 56.9 | 43.0 | 15.8 | 50.9 |
| Morsy-1953 | 60.0 | 30.0 | 49.4 | 59.0 | 22.1 | 55.3 | 48.0 | 26.5 | 45.0 | 54.1 | 20.2 | 43.7 |
| Morsy-1954 | 29.5 | 20.8 | 55.0 | 29.0 | 15.9 | 26.7 | 53.6 | 17.5 | 51.5 | 21.7 | 10.2 | 50.5 |
| Morsy-1955 | 58.1 | 9.1 | 49.7 | 55.1 | 2.0 | 52.8 | 46.0 | 6.1 | 45.4 | 50.0 | 1.9 | 39.9 |
| Morsy-1956 | 50.0 | 13.5 | 67.0 | 44.2 | 9.3 | 45.1 | 67.0 | 12.2 | 46.0 | 41.0 | 7.0 | 66.0 |
| Morsy-1957 | 22.9 | 46.0 | 61.5 | 19.7 | 43.2 | 18.0 | 59.9 | 48.8 | 59.2 | 13.5 | 42.5 | 56.5 |
| Morsy-1958 | 20.8 | 18.5 | 49.6 | 17.0 | 14.9 | 15.4 | 48.0 | 16.0 | 47.0 | 13.0 | 11.6 | 34.3 |
| Morsy-1959 | 55.2 | 10.0 | 50.8 | 51.9 | 31.0 | 51.6 | 46.9 | 37.3 | 46.4 | 50.6 | 30.0 | 45.1 |
| Morsy-1960 | 47.0 | 40.7 | 30.2 | 45.0 | 34.8 | 40.9 | 30.0 | 37.2 | 26.8 | 37.4 | 31.7 | 25.0 |
| Morsy-1961 | 21.6 | 21.0 | 19.0 | 18.0 | 35.7 | 16.2 | 17.5 | 40.0 | 15.0 | 13.8 | 34.1 | 19.7 |
| Morsy-1962 | 47.3 | 39.0 | 60.0 | 44.5 | 30.8 | 40.9 | 66.8 | 35.4 | 54.3 | 38.6 | 30.0 | 54.1 |
| Morsy-1963 | 50.8 | 40.2 | 58.5 | 47.9 | 35.6 | 43.5 | 55.0 | 39.0 | 56.4 | 40.0 | 33.5 | 52.8 |
| Morsy-1964 | 41.2 | 50.0 | 60.0 | 46.3 | 44.5 | 37.3 | 45.9 | 48.5 | 60.0 | 36.1 | 43.0 | 60.0 |
| Morsy-1965 | 25.0 | 20.1 | 61.5 | 23.4 | 15.0 | 20.1 | 59.4 | 19.0 | 54.1 | 17.5 | 11.7 | 51.9 |
| Morsy-1966 | 42.7 | 40.0 | 56.0 | 41.0 | 36.2 | 37.0 | 55.0 | 37.1 | 652.5 | 35.3 | 35.8 | 50.0 |
| Morsy-1967 | 50.3 | 40.6 | 57.4 | 48.5 | 34.9 | 45.7 | 54.0 | 40.0 | 54.0 | 40.8 | 30.3 | 51.7 |
| Morsy-1968 | 58.0 | 41.8 | 30.0 | 56.0 | 37.0 | 51.0 | 37.1 | 38.5 | 33.2 | 47.4 | 36.0 | 32.5 |
| Morsy-1969 | 50.2 | 50.0 | 49.0 | 49.1 | 46.1 | 43.6 | 45.7 | 48.0 | 41.6 | 40.6 | 42.8 | 35.1 |
| Morsy-1970 | 38.5 | 39.4 | 62.8 | 36.2 | 33.5 | 30.3 | 66.5 | 35.8 | 61.0 | 30.0 | 32.0 | 58.6 |
| Morsy-1971 | 47.4 | 40.0 | 53.4 | 45.0 | 36.4 | 44.0 | 49.9 | 36.9 | 47.5 | 43.2 | 35.4 | 47.0 |
| Morsy-1972 | 30.0 | 45.8 | 28.1 | 33.0 | 37.8 | 25.0 | 87.2 | 41.9 | 84.0 | 22.0 | 35.9 | 20.0 |
| Morsy-1973 | 22.8 | 44.0 | 60.2 | 18.0 | 40.0 | 16.6 | 66.0 | 41.7 | 65.3 | 13.9 | 38.6 | 60.3 |
| Morsy-1974 | 53.5 | 38.2 | 20.0 | 46.7 | 31.9 | 48.5 | 14.9 | 35.2 | 12.7 | 44.0 | 28.7 | 10.9 |
| Morsy-1975 | 49.8 | 53.0 | 65.0 | 50.0 | 50.0 | 43.9 | 65.3 | 52.0 | 50.4 | 42.0 | 49.4 | 55.8 |
| Morsy-1976 | 38.4 | 42.0 | 58.3 | 33.5 | 36.8 | 30.4 | 54.1 | 39.5 | 51.6 | 26.3 | 35.0 | 47.0 |
| Morsy-1977 | 51.5 | 53.0 | 25.0 | 47.4 | 48.0 | 46.6 | 22.5 | 50.6 | 15.9 | 43.4 | 44.5 | 12.2 |
| Morsy-1978 | 20.0 | 30.0 | 60.4 | 18.9 | 24.2 | 14.8 | 66.0 | 28.0 | 52.1 | 14.0 | 21.2 | 45.5 |
| Morsy-1979 | 54.5 | 20.8 | 49.0 | 54.1 | 15.3 | 47.1 | 47.3 | 17.2 | 44.0 | 42.5 | 15.0 | 38.4 |
| Morsy-1980 | 20.3 | 19.0 | 44.0 | 17.1 | 15.0 | 16.5 | 41.6 | 18.0 | 38.7 | 13.8 | 13.7 | 36.0 |
| Morsy-1981 | 33.9 | 22.7 | 60.5 | 30.0 | 18.4 | 30.0 | 56.0 | 19.9 | 52.0 | 27.5 | 16.6 | 50.3 |
| Morsy-1982 | 54.2 | 40.4 | 34.0 | 50.7 | 34.9 | 46.1 | 31.2 | 37.5 | 30.3 | 45.2 | 34.0 | 30.0 |
| Morsy-1983 | 19.1 | 14.0 | 67.1 | 16.8 | 10.2 | 14.3 | 64.0 | 11.8 | 64.5 | 11.2 | 9.1 | 60.6 |
| Morsy-1984 | 47.6 | 39.1 | 14.0 | 46.4 | 32.8 | 40.5 | 11.1 | 35.4 | 10.3 | 38.1 | 29.5 | 9.5 |
| Morsy-1985 | 50.8 | 40.5 | 50.2 | 50.0 | 32.9 | 64.9 | 48.0 | 37.5 | 44.0 | 45.4 | 29.0 | 41.9 |
| Morsy-1986 | 50.5 | 10.7 | 61.3 | 47.9 | 5.1 | 44.2 | 50.0 | 10.0 | 37.2 | 20.7 | 1.8 | 25.4 |
| Morsy-1987 | 37.0 | 19.0 | 50.0 | 35.5 | 16.5 | 34.0 | 41.9 | 19.0 | 40.5 | 33.0 | 17.2 | 32.6 |
| Morsy-1988 | 53.5 | 30.0 | 56.3 | 51.0 | 23.0 | 49.6 | 52.4 | 26.0 | 42.7 | 45.3 | 21.6 | 39.3 |
| Morsy-1989 | 49.6 | 9.9 | 60.0 | 45.9 | 5.0 | 40.4 | 59.1 | 7.3 | 65.9 | 40.0 | 2.5 | 65.1 |
| Morsy-1990 | 32.4 | 35.0 | 60.6 | 30.0 | 29.0 | 28.0 | 66.7 | 33.5 | 66.3 | 25.8 | 27.4 | 60.0 |
| Morsy-1991 | 36.0 | 20.2 | 60.5 | 32.7 | 12.0 | 31.5 | 57.5 | 17.8 | 53.0 | 29.0 | 10.5 | 50.2 |
| Morsy-1992 | 51.9 | 50.0 | 49.0 | 50.0 | 40.9 | 44.7 | 45.0 | 45.0 | 41.7 | 54.0 | 39.7 | 39.2 |
| Morsy-1993 | 54.5 | 44.5 | 30.0 | 52.7 | 39.5 | 52.0 | 26.2 | 43.1 | 23.1 | 50.1 | 38.0 | 20.0 |
| Morsy-1994 | 13.0 | 40.4 | 57.7 | 11.6 | 40.0 | 8.9 | 52.3 | 40.2 | 50.4 | 7.9 | 39.1 | 50.0 |
| Morsy-1995 | 55.0 | 38.9 | 60.0 | 41.8 | 32.7 | 40.8 | 57.0 | 36.2 | 57.0 | 39.3 | 31.0 | 55.5 |
| Morsy-1996 | 44.6 | 22.0 | 61.3 | 44.0 | 20.0 | 42.1 | 68.4 | 21.3 | 65.6 | 39.5 | 19.3 | 58.1 |
| Morsy-1997 | 60.0 | 50.3 | 30.0 | 55.9 | 44.6 | 50.2 | 30.0 | 47.1 | 41.0 | 47.8 | 42.8 | 35.2 |
| Morsy-1998 | 19.1 | 20.7 | 65.3 | 16.3 | 13.5 | 15.0 | 71.5 | 16.5 | 59.5 | 14.6 | 12.8 | 59.0 |
| Morsy-1999 | 44.9 | 10.0 | 19.4 | 43.0 | 7.8 | 46.9 | 25.8 | 18.0 | 15.2 | 46.5 | 16.5 | 22.9 |
| Morsy-2000 | 47.1 | 40.6 | 50.0 | 47.0 | 35.7 | 46.3 | 44.5 | 37.4 | 42.8 | 45.0 | 33.2 | 41.6 |
| Morsy-2001 | 59.5 | 47.0 | 50.0 | 55.0 | 40.3 | 53.0 | 39.0 | 45.0 | 41.0 | 52.2 | 39.5 | 37.5 |
| Morsy-2002 | 45.0 | 30.3 | 41.0 | 44.0 | 22.0 | 40.8 | 33.0 | 25.5 | 35.6 | 40.0 | 20.9 | 32.8 |
| Morsy-2003 | 50.0 | 22.6 | 50.4 | 47.3 | 18.5 | 45.6 | 47.1 | 20.3 | 39.3 | 43.6 | 18.0 | 38.4 |
| Morsy-2004 | 90.0 | 20.0 | 28.5 | 88.1 | 15.0 | 80.2 | 25.5 | 27.2 | 25.4 | 80.0 | 12.6 | 80.0 |
| Morsy-2005 | 25.4 | 45.3 | 40.6 | 24.0 | 40.1 | 27.9 | 37.9 | 43.0 | 32.0 | 25.9 | 39.4 | 27.1 |
| Morsy-2006 | 60.0 | 48.0 | 35.2 | 58.0 | 44.5 | 55.5 | 33.0 | 46.1 | 32.5 | 50.6 | 43.1 | 30.5 |
| Morsy-2007 | 15.3 | 39.6 | 57.0 | 11.9 | 36.0 | 11.0 | 53.2 | 39.0 | 62.4 | 10.3 | 37.8 | 55.0 |
| Morsy-2008 | 18.0 | 14.2 | 64.2 | 18.0 | 9.2 | 16.5 | 59.0 | 11.5 | 69.2 | 16.0 | 8.0 | 66.7 |
| Morsy-2009 | 35.0 | 50.0 | 23.5 | 32.7 | 35.0 | 29.9 | 25.1 | 37.9 | 29.6 | 30.5 | 32.9 | 24.7 |
| Morsy-2010 | 9.9 | 20.5 | 20.5 | 7.9 | 16.0 | 4.7 | 17.2 | 17.6 | 17.0 | 10.9 | 14.5 | 15.8 |
| Morsy-2011 | 58.0 | 20.0 | 41.3 | 56.0 | 15.2 | 55.4 | 37.4 | 17.3 | 26.4 | 52.6 | 15.0 | 29.0 |
| Morsy-2012 | 39.6 | 40.7 | 64.0 | 37.1 | 35.8 | 33.2 | 60.0 | 37.0 | 60.0 | 33.0 | 33.8 | 60.0 |
| Morsy-2013 | 52.1 | 50.1 | 30.5 | 40.0 | 42.4 | 44.0 | 45.0 | 45.9 | 38.0 | 39.3 | 41.7 | 41.6 |
| Morsy-2014 | 50.5 | 59.4 | 62.8 | 60.65 | 68.19 | 69.19 | 51.81 | 86.2 | 41.6 | 62.92 | 77.64 | 32.0 |
| Morsy-2015 | 50.4 | 20.0 | 50.5 | 50.0 | 17.0 | 48.5 | 42.9 | 18.2 | 46.8 | 75.0 | 15.6 | 40.0 |
| Morsy-2016 | 55.0 | 49.3 | 47.0 | 49.4 | 42.5 | 48.0 | 45.1 | 47.5 | 40.5 | 47.2 | 41.5 | 40.0 |
Although real wastewater treatment systems often have to deal with a mixture of heavy metals, most research work still focuses on the sorption capacity under a single metal system. As is evident from Table 2, the metal biosorption capacity of Actinomycete isolates under binary and ternary systems was observed to exhibit no significant interference consequent to competition between metals for bioremoval. In case of the isolate MORSY1948, when in the presence of a binary system composed of Ni2++Zn2+ or Cr6++Zn2+, it was observed that the presence of Zn2+ led to an enhancement of the biosorption capacity of Ni2+ and Cr6+ by 20.6% and 15.9%, respectively. This observation was in parallel with results that suggested an increase in the removal efficiency of Zn2+, in these bimetallic systems, by 5.8% and 12.2%, respectively. Both these results are suggestive of lesser observable interference between these three metals for binding to the metal binding sites on the biomass of MORSY1948 (Table 2). Similarly, it was observed that in a binary system composed of Cr6+ and Ni2+, the removal capacity of MORSY1948 was estimated to be 51.0% and 79.5% as compared to 47.4% and 60.1% in single system, respectively. In case of the second strain MORSY2014, Zn2+ removal was observed to decline by 17.5% over its maximum removal efficiency of 62.8% when present in a binary system along with Ni2+. In contrast, a significant 37% enhancement in Ni2+ removal was observed in the presence of Zn2+ as secondary metal. When the dead biomass of MORSY2014 was used as a biosorbent, it was also observed that the presence of Zn2+ enhanced the removal of Cr6+ in bimetallic system composed of Cr6++Zn2+ (59.4%–86.2%) but Cr6+ suppressed Zn2+ removal efficiency to 41.6%. Interestingly, in the bimetallic system of Cr2++Ni2+ there was no observable competition between the biosorption capacities of both metal ions as the removal capability for both metals were seen to increase by 14.8% and 20.1%, respectively (Table 2).
In a multi-metallic ternary mixture composed of Ni2++Cr6++Zn2+ in aqueous solution, it was observed that the removal efficiency the MORSY1948 biomass increased from 60.1%, 47.4% and 67.4% to 74%, 58.4% and 81.3%, respectively (Table 2). Similarly, in case of MORSY2014 the removal efficiency for Ni2+ and Cr6+ was observed to significantly increase by 24.6% and 30.7%, respectively when observed in a ternary system. However the same cannot be said for Zn2+ which witnessed a sharp decrease from 62.8% in the single metal system to 32.0% in the ternary metal system. This observation can be hypothetically explained as a consequence result of competitive interactions for metal binding sites and accumulation of Zn2+ as well as the other two metals, Ni2+ and Cr6+, inside the MORSY2014 biomass (Table 2). Very limited literature is available regarding sorption of metals from heavy metal mixtures. A published study by Kaewsam40 reported that in mixed metal systems, Cu2+ uptake was significantly affected by presence of other heavy metals such as Ag+, Mn2+, Co2+, Ni2+, Fe2+, Cd2+ and Pb2+. A report by Rho and Kim5 illustrated the order of adsorption potential of different heavy metals, in case of S. viridochromogenes was Zn2+>Cu2+>Pb2+>Cd2+ in single and mixed metal reactions, whereas the same for S. chromofuscus K101 was Zn2+>Pb2+>Fe2+≥Cu2+≥Cd2+.7
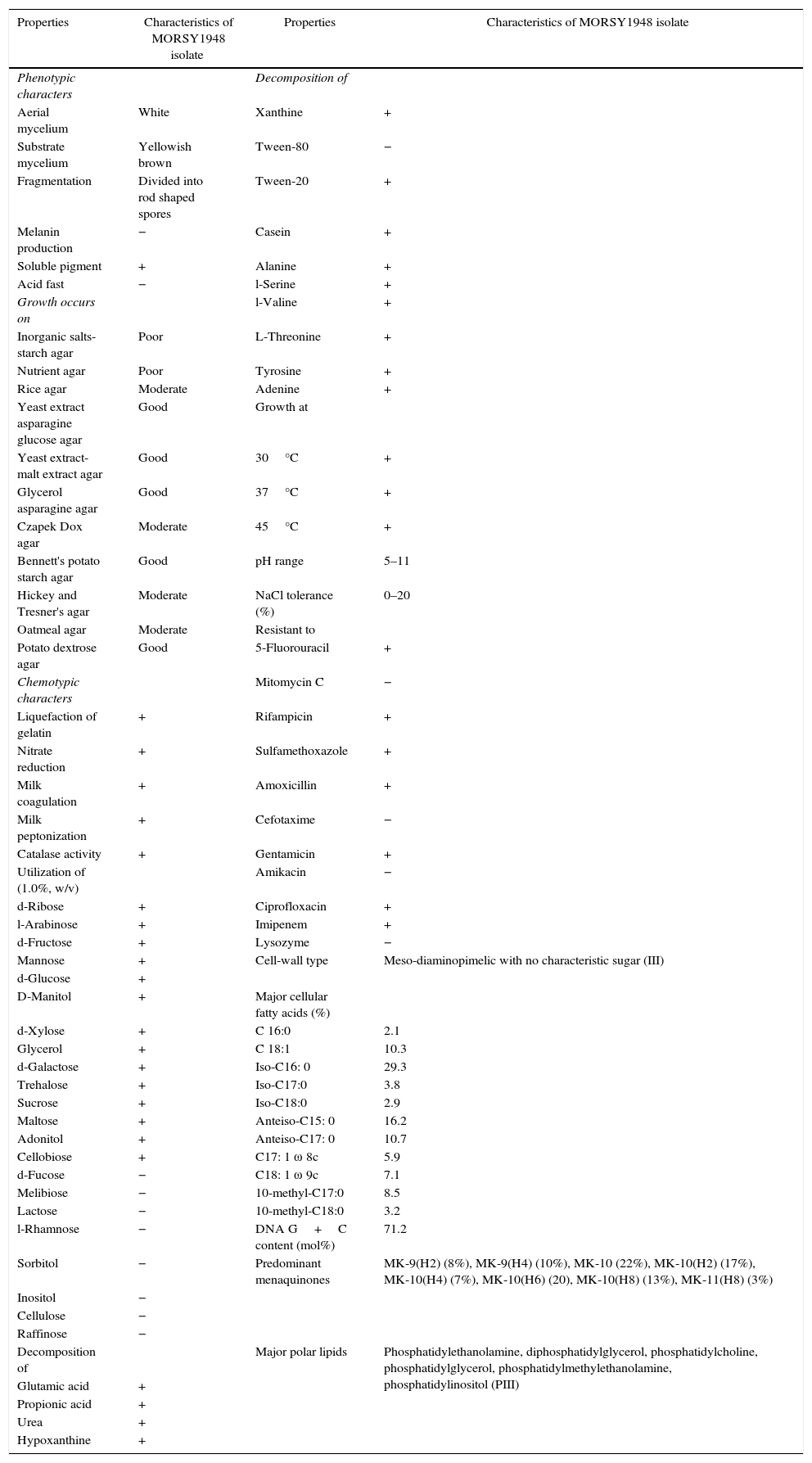
Identification the high biosorption potential MORSY1948 and MORSY2014 isolatesCharacteristics of the MORSY1948 strainThe phenotypic and chemotaxonomic characteristics of MORSY1948 strain, as summarized in Table 3, were found to be consistent with those described for the genus Nocardiopsis. The mature vegetative hyphae were long, well-developed and fragmented into rod- shaped elements. The strain displayed a white aerial mycelium which was observed to turn into a yellowish-white shade in older cultures. Also, the substrate mycelium was yellowish brown in color which matched with that observed for most of the Nocardiopsis species. Soluble pigments were produced but melanin formation was not observed. Growth patterns observed can be classified as good in case of yeast extract-malt extract agar, Bennett's potato starch agar, yeast extract asparagine glucose agar, glycerol asparagine agar and potato dextrose agar; moderate growth was seen on oatmeal agar, rice agar, Czapek Dox agar and Hickey and Tresner's agar and poor growth was seen on inorganic salts-starch agar and nutrient agar (Table 3). Chemotaxonomic characteristics of strain MORSY1948 were consistent with its classification as a member of the genus Nocardiopsis. Tests such as the liquefaction of gelatin, catalase activity, nitrate reduction, milk coagulation and peptonization were positive while the acid fast test was negative. MORSY1948 strain was capable of utilizing ribose, arabinose, fructose, mannose, glucose, mannitol, xylose, glycerol, galactose, trehalose, sucrose, maltose, adonitol, and cellobiose as a sole carbon source but not fucose, melibiose, lactose, rhamnose, sorbitol, inositol, cellulose, and raffinose. Besides this, it was unable to degrade Tween 80 (Table 3). Strain MORSY1948 exhibited robust growth under the following range of conditions: temperature – 30, 37 and 45°C, pH – between 5 and 11, and NaCl concentrations as high as 20% (w/v). The strain was susceptible to antimicrobial agents such as mitomycin C, cefotaxime, amikacin, and lysozyme (Table 3). The whole-cell hydrolysates were found to contain meso-diaminopimelic acid as the only peptidoglycan diamino acid without a characteristic sugar (type III). Polar lipid pattern revealed the presence of the diagnostic phosphatidyl ethanolamine (PE), diphosphatidyl glycerol (DPG), phosphatidyl choline (PC), phosphatidyl glycerol (PG), phosphatidyl methylethanolamine (PME), and phosphatidyl inositol (PI) (pattern III12). This phospholipid pattern is known to be found in the species of the genera Nocardiopsis, Actinopolyspora, Saccharopolyspora, and Pseudonocardia. Nocardiopsis strains, however, can easily be differentiated from these taxa by the occurrence of PME, presence of high amounts of PG, and the lack of hydroxy-phosphatidyl ethanolamine. The predominant menaquinones found were: MK-9(H2) (8%), MK-9(H4) (10%), MK-10 (22%), MK-10(H2) (17%), MK-10(H4) (7%), MK-10(H6) (20%), MK-10(H8) (13%) and MK-11(H8) (3%). The combination of fatty acids in this strain is unique among Nocardiopsis species16,20 as it is composed of C 16:0 (2.1%), C 18:1(10.3%), iso-C16: 0 (29.3%), iso-C17:0 (3.8%), iso-C18:0 (2.9%), anteiso-C15: 0 (16.2%), anteiso-C17: 0 (10.7%), C17: 1 ω 8c (5.9%), C18: 1 ω 9c (7.1%), 10-methyl-C17:0 (8.5%) and 10-methyl-C18:0 (3.2%) (fatty acid type 3d). The high amount of anteiso-fatty acids in combination with 10-methyl-branched fatty acids (fatty acid type 3d) is diagnostic of species belonging to the genus Nocardiopsis. Moreover, the DNA G+C content was equivalent to 71.2mol% (Table 3). The members of the genus Nocardiopsis are known to produce bioactive metabolites such as griseusin D, apoptolidin, methylpendolmycin, thiopeptide and naphthospironone A.41
Phenotypic and chemotaxonomic characteristics of MORSY1948 isolate.
| Properties | Characteristics of MORSY1948 isolate | Properties | Characteristics of MORSY1948 isolate |
|---|---|---|---|
| Phenotypic characters | Decomposition of | ||
| Aerial mycelium | White | Xanthine | + |
| Substrate mycelium | Yellowish brown | Tween-80 | − |
| Fragmentation | Divided into rod shaped spores | Tween-20 | + |
| Melanin production | − | Casein | + |
| Soluble pigment | + | Alanine | + |
| Acid fast | − | l-Serine | + |
| Growth occurs on | l-Valine | + | |
| Inorganic salts-starch agar | Poor | L-Threonine | + |
| Nutrient agar | Poor | Tyrosine | + |
| Rice agar | Moderate | Adenine | + |
| Yeast extract asparagine glucose agar | Good | Growth at | |
| Yeast extract-malt extract agar | Good | 30°C | + |
| Glycerol asparagine agar | Good | 37°C | + |
| Czapek Dox agar | Moderate | 45°C | + |
| Bennett's potato starch agar | Good | pH range | 5–11 |
| Hickey and Tresner's agar | Moderate | NaCl tolerance (%) | 0–20 |
| Oatmeal agar | Moderate | Resistant to | |
| Potato dextrose agar | Good | 5-Fluorouracil | + |
| Chemotypic characters | Mitomycin C | − | |
| Liquefaction of gelatin | + | Rifampicin | + |
| Nitrate reduction | + | Sulfamethoxazole | + |
| Milk coagulation | + | Amoxicillin | + |
| Milk peptonization | + | Cefotaxime | − |
| Catalase activity | + | Gentamicin | + |
| Utilization of (1.0%, w/v) | Amikacin | − | |
| d-Ribose | + | Ciprofloxacin | + |
| l-Arabinose | + | Imipenem | + |
| d-Fructose | + | Lysozyme | − |
| Mannose | + | Cell-wall type | Meso-diaminopimelic with no characteristic sugar (III) |
| d-Glucose | + | ||
| D-Manitol | + | Major cellular fatty acids (%) | |
| d-Xylose | + | C 16:0 | 2.1 |
| Glycerol | + | C 18:1 | 10.3 |
| d-Galactose | + | Iso-C16: 0 | 29.3 |
| Trehalose | + | Iso-C17:0 | 3.8 |
| Sucrose | + | Iso-C18:0 | 2.9 |
| Maltose | + | Anteiso-C15: 0 | 16.2 |
| Adonitol | + | Anteiso-C17: 0 | 10.7 |
| Cellobiose | + | C17: 1 ω 8c | 5.9 |
| d-Fucose | − | C18: 1 ω 9c | 7.1 |
| Melibiose | − | 10-methyl-C17:0 | 8.5 |
| Lactose | − | 10-methyl-C18:0 | 3.2 |
| l-Rhamnose | − | DNA G+C content (mol%) | 71.2 |
| Sorbitol | − | Predominant menaquinones | MK-9(H2) (8%), MK-9(H4) (10%), MK-10 (22%), MK-10(H2) (17%), MK-10(H4) (7%), MK-10(H6) (20), MK-10(H8) (13%), MK-11(H8) (3%) |
| Inositol | − | ||
| Cellulose | − | ||
| Raffinose | − | ||
| Decomposition of | Major polar lipids | Phosphatidylethanolamine, diphosphatidylglycerol, phosphatidylcholine, phosphatidylglycerol, phosphatidylmethylethanolamine, phosphatidylinositol (PIII) | |
| Glutamic acid | + | ||
| Propionic acid | + | ||
| Urea | + | ||
| Hypoxanthine | + |
The MORSY2014 strain was subjected to a broad range of phenotypic and chemotaxonomic analysis. The strain was characterized as strictly aerobic, gram positive and slightly acid-fast. Pale white aerial mycelium with well-developed yellowish orange substrate mycelium that had extensively irregular branched hyphae with a tendency to fragmentation into rods and coccoid elements was observed. Other characteristics noted were: growth on agar, filamentous margins, beaded appearance, and soluble pigments formation (Table 4). The strain exhibited poor growth on oatmeal agar, moderate growth on inorganic salts-starch agar, rice agar and Czapek Dox agar and good growth on yeast extract asparagine glucose agar, Bennett's potato starch agar, Hickey and Tresner's agar and potato dextrose agar. Although the strain was positive for catalase, urease, and β-galactosidase activity, but was negative for arylsulfatase activity. The strain was able to utilize d-ribose, melibiose, d-fructose, mannose, d-glucose, d-mannitol, glycerol, d-galactose, trehalose, lactose, sorbitol, inositol, adonitol and salicin but not l-arabinose, d-fucose, d-xylose, l-rhamnose, sucrose, maltose and raffinose. Casein, gluconic acid, propionic acid, uric acid, hypoxanthine, Tween 80, Tween 20, l-valine, tyrosine, and urea were observed to be utilized by the MORSY2014 strain but the same was not observed for ornithine, adenine, and xanthine. Growth on Bennett agar was observed at 25°C as well as at 37°C and also at pH 10 but no growth was observed at 45°C or at pH 12 (Table 4). MORSY2014 strain exhibited susceptibility to antimicrobial agents such as mitomycin C, rifampicin, cefotaxime, amikacin, ciprofloxacin, and imipenem. The G+C content of the genomic DNA was 63.9mol%. The fatty acid pattern detected in MORSY2014 strain was as follows (%): C13: 0 (3.22), C14: 0 (10.71), C15: 0 (1.15), C16: 0 (24.25), C16: 1 (16.11), C17: 0 (1.22), C18: 1 (6.34), 10-methyl C18: 0 (6.35), C16: 1 ω 9c (1.25), C17: 1 ω8c (1.19), C18: 1 ω9c (6.20), C16: 1 ω 7c (1.73) and C16: 1cis (20.28). Whole-cell hydrolysates were found to contain meso-diaminopimelic acid with arabinose and galactose as characteristic sugars (type IV). The Nocardia-specific quinones were cyclo MK-8(H4), which was found to represent 78.26% of the all menaquinones detected in MORSY2014; lesser amounts of MK-8(H2) (15.47%), MK-8(H4) (5.12%), and MK-8(H6) (1.15%) were also seen. Major polar lipids observed were: phosphatidylethanolamine, phosphatidylinositol, diphosphatidylglycerol and phosphatidylinositol mannoside (Table 4). Hence, as it is evident from the above that several physiological and biochemical characteristics support the assignment of strain MORSY2014 to the genus Nocardia.
phenotypic and chemotypic characteristics of MORSY2014 isolate.
| Properties | Characteristics of MORSY2014 isolate | Properties | Characteristics of MORSY2014 isolate |
|---|---|---|---|
| Phenotypic characters | Decomposition of (0.5%, w/v) | ||
| Aerial mycelium | Pale white | Hypoxanthine | + |
| Substrate mycelium | Yellowish orange | Xanthine | − |
| Filamentous margins | + | Tween-80 | + |
| Growth into agar | + | Tween-20 | + |
| Rough and waxy colony | + | l-Valine | + |
| Soluble pigment | + | Ornithine | − |
| Fragmentation into rods and coccoid elements | + | Tyrosine | + |
| Beaded | + | Urea | + |
| Acid fast | + | Adenine | − |
| Growth occurs on | Growth at (Bennett's agar) | ||
| Inorganic salts-starch agar | Moderate | 30°C | + |
| Rice agar | Moderate | 37°C | + |
| Yeast extract asparagine glucose agar | Good | 45°C | − |
| Czapek Dox agar | Moderate | pH 10 | + |
| Bennett's potato starch agar | Good | pH 12 | − |
| Hickey and Tresner's agar | Good | NaCl tolerance (%) | Up to 18 |
| Oatmeal agar | Poor | Resistant to | |
| Potato dextrose agar | Good | 5-Fluorouracil | + |
| Chemotypic characters | Mitomycin C | − | |
| Liquefaction of gelatin | + | Rifampicin | − |
| Nitrate reduction | + | Sulfamethoxazole | + |
| Catalase | + | Amoxicillin | + |
| Urease | + | Cefotaxime | − |
| β-Galactosidase | + | Gentamicin | + |
| Arylsulfatase | − | Amikacin | − |
| Utilization of (1.0%, w/v) | Ciprofloxacin | − | |
| d-Ribose | + | Imipenem | − |
| l-Arabinose | − | Lysozyme | + |
| d-Fucose | − | Major fatty acids (%) | |
| Melibiose | + | C13: 0 | 3.22 |
| d-Fructose | + | C14: 0 | 10.71 |
| Mannose | + | C15: 0 | 1.15 |
| d-Glucose | + | C16: 0 | 24.25 |
| D-Manitol | + | C16: 1 | 16.11 |
| d-Xylose | − | C17: 0 | 1.22 |
| Glycerol | + | C18: 1 | 6.34 |
| d-Galactose | + | 10-methyl C18: 0 | 6.35 |
| Trehalose | + | C16: 1 ω 9c | 1.25 |
| Lactose | + | C17: 1 ω8c | 1.19 |
| l-Rhamnose | − | C18: 1 ω9c | 6.20 |
| Sucrose | − | C16: 1 ω 7c | 1.73 |
| Maltose | − | C16:1cis | 20.28 |
| Sorbitol | + | Cell wall type | Meso-diaminopimelic with arabinose and galactose as characteristic sugars (IV) |
| Inositol | + | ||
| Adonitol | + | G+C (mol%) | 63.9 |
| Salicin | + | Major quinones | Cyclo MK-8(H4) (78.26), MK-8(H2) (15.47%), MK-8(H4) (5.12%) and MK-8(H6) (1.15%) |
| Raffinose | − | ||
| Decomposition of (0.5%, w/v) | Major Polar lipids | Phosphatidylethanolamine, Phosphatidylinositol, diphosphatidylglycerol, phosphatidylinositol mannoside | |
| Casein | + | ||
| Gluconic acid | + | ||
| Propionic acid | + | ||
| Uric acid | + |
The almost-complete 16S rDNA sequences of strains MORSY1948 and MORSY2014 (accession numbers KP979750 and KP979751, respectively) were compared to the sequences of members of the order Actinomycetales. It was observed that the members of the genus Nocardiopsis and Nocardia, respectively, were the closest phylogenetic neighbors. The values were seen to range between 96% (Nocardiopsis dassonvillei Subsp. albirubida VTT E-062983) and 98% (Nocardiopsis sp. TRM46486) for isolate MORSY1948 to 98% (Nocardia sp. OAct 132) and 99% (N. cummidelens DSM 44490) for isolate MORSY2014 (Fig. 1). Based on the 16S rDNA analyses and phylogenetic data, it was concluded that both the isolates, MORSY1948 and MORSY2014, merit species status within the genus Nocardiopsis and Nocardia, respectively. Isolates MORSY1948 and MORSY2014 can be differentiated from the Nocardiopsis and Nocardia species by a combination of morphological, physiological, chemotaxonomic and 16S rDNA analyses data. Based on these results, it was concluded that strains MORSY1948 and MORSY2014 are species of the genus Nocardiopsis and Nocardia; and hence were given the names as Nocardiopsis sp. MORSY1948 and Nocardia sp. MORSY2014.
Factors that affect the biosorption process of heavy metals by live and dead biomasses of Nocardiopsis sp. MORSY1948, and Nocardia sp. MORSY2014 strainsEffect of different temperatures
An analysis of the strains Nocardiopsis sp. MORSY1948 and Nocardia sp. MORSY2014 for reduction efficiency at different temperatures revealed that irrespective of growth temperature, reduction efficiency for all heavy metals under study was higher for dead biomass rather than it was for live (Fig. 2a and b). Bioremoval capacity of Ni2+, Cr6+ and Zn2+ was increased by 60.5%, 47.5% and 67.8% when non-active cells of Nocardiopsis sp. MORSY1948 were incubated at 40°C as compared to their live counterparts (Fig. 2a). The removal efficiency at 40°C for the dead biomass of Nocardiopsis sp. MORSY2014 for Ni2+, Cr6+ and Zn2+ was 1.45 fold, 2.11 fold, and 1.55 fold higher than that of the live biomass (Fig. 2b). Although living biomass has an additional capacity for heavy metal biosorption as a result of metabolic entrapment, nonliving biomasses have several advantages to offer. These include their ease of treatment, their strong affinity for metal ions because of the lack of a proton production mechanism during metabolism and no metal toxicity issues of the type that can result in cell death in live cells. Additionally, the cultivation of live biomass requires supplementation with nutrients, which can increase the biological and chemical oxygen demands on the treated water.7 Our results are line with that reported by Simeonova et al.,31 Al Turk and Kiki4 and Daboor et al.7 in which the authors claim that maximum biosorption of heavy metals was obtained when dead biomasses of S. fradiae, halophilic Actinomycetes and S. chromofuscus K101 were used for the purpose. Interestingly, it was observed that when 100mgL−1 solutions of metal ions, Ni2+, Cr6+ and Zn2+, were individually applied onto 0.3% biomass of MORSY1948 or MORSY2014, temperature was discovered to function as a significant parameter influencing biosorption. This indicates that the biosorption process is endothermic in nature.
 Effect of different temperatures on the biosorption capacity of Ni2+, Cr6+ and Zn2+ (%) from aquous solution by the live and dead cells of Nocardiopsis sp. MORSY1948. (b) Effect of different temperatures on the biosorption capacity of Ni2+, Cr6+ and Zn2+ (%) from aquous solution by the live and dead cells of Nocardia sp. MORSY2014.")
(a) Effect of different temperatures on the biosorption capacity of Ni2+, Cr6+ and Zn2+ (%) from aquous solution by the live and dead cells of Nocardiopsis sp. MORSY1948. (b) Effect of different temperatures on the biosorption capacity of Ni2+, Cr6+ and Zn2+ (%) from aquous solution by the live and dead cells of Nocardia sp. MORSY2014.
By increasing the temperature from 30°C to 40°C, the removal of Ni2+, Cr6+ and Zn2+ by Nocardiopsis sp. MORSY1948 strain was elevated from 22.5%, 17.3% and 19.7% to 39.4%, 28.0%, 37.9%, respectively (live biomass) and from 49.0%, 28.5% and 37.2% to 60.5%, 47.5% and 67.8%, respectively (dead biomass) (Fig. 2a). For Nocardia sp. MORSY2014 (live biomass), the potent sorption percentage for Ni2+, Cr6+ and Zn2+ at 50°C was determined as 49.2%, 38.5% and 50.2% respectively as compared to 14%, 15% and 23.6% at 30°C. The same for the dead biomass was estimated to be 80.0%, 75.4% and 76.9% respectively at 50°C as compared to 24.1%, 20.9% and 39.4% at 30°C (Fig. 2b). The increase in adsorption percentage with elevation in temperature can be attributed to the several factors such as a change in the pore size of the adsorbent leading to a greater inter-particle diffusion within the pores, the creation of new active sites on the sorbent, a temperature-based acceleration of some slow adsorption steps, an enhancement in the mobility of metal ions from the bulk solution toward the adsorbent surface, and/or an enhancement in the chemical affinity of the metal cations for the surface of adsorbent.42
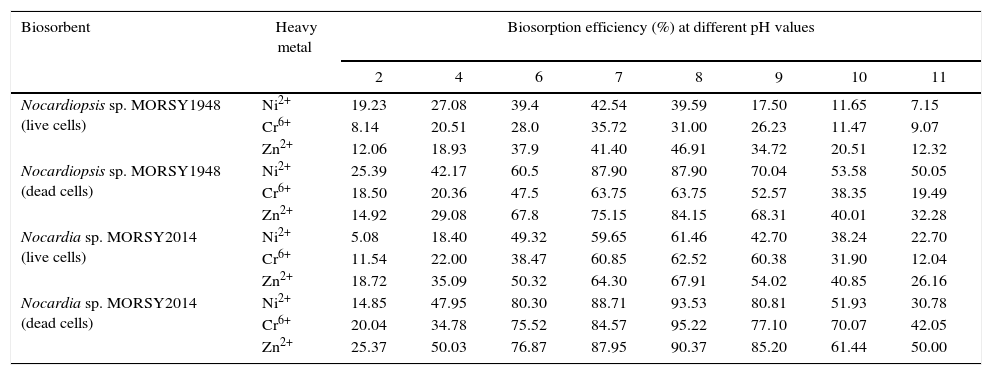
Effect of different pH valuesBy increasing pH from 2 to 7, absorption capability of the live biomass of Nocardiopsis sp. MORSY1948 toward Ni2+ and Cr6+ was increased 2.21 fold and 4.39 fold respectively, while at pH 8, Zn2+ bioremoval efficiency increased 3.89 fold. It was noticed that the dead biomass removed 87.9% and 63.75% of Ni2+ and Cr6+ at pH 7, respectively, and 84.15% of Zn2+ was removed at pH 8 (Table 5). In case of Nocardia sp. MORSY2014, only 61.46% (Ni2+), 62.52% (Cr6+) and 67.91% (Zn2+) removal potential was achieved by the live biomass of the strain which is in sharp contrast to the much superior removal efficiency of the dead biomass at pH 8 [93.53% (Ni2+), 95.22% (Cr2+) and 90.37% (Zn2+)] (Table 5). The biosorption capacity of both strains was observed to significantly decrease at acidic pH values. This is hypothesized to be due to the fact that H+ ions compete with metal ions for the negatively charged binding sites thereby hindering them from reaching the binding sites of the adsorbent (repulsive forces). However, at pH 4 and above, an ion exchange mechanism occurs via H+ ion and the negatively charged groups of biomass result in a drastic increase in the heavy metal removal efficiency.30
Effect of pH on biosorption efficiency (%) of the heavy metals Ni2+, Cr6+ and Zn2+ by live and dead biomass of Nocardiopsis sp. MORSY1948 and Nocardia sp. MORSY2014.
| Biosorbent | Heavy metal | Biosorption efficiency (%) at different pH values | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 2 | 4 | 6 | 7 | 8 | 9 | 10 | 11 | ||
| Nocardiopsis sp. MORSY1948 (live cells) | Ni2+ | 19.23 | 27.08 | 39.4 | 42.54 | 39.59 | 17.50 | 11.65 | 7.15 |
| Cr6+ | 8.14 | 20.51 | 28.0 | 35.72 | 31.00 | 26.23 | 11.47 | 9.07 | |
| Zn2+ | 12.06 | 18.93 | 37.9 | 41.40 | 46.91 | 34.72 | 20.51 | 12.32 | |
| Nocardiopsis sp. MORSY1948 (dead cells) | Ni2+ | 25.39 | 42.17 | 60.5 | 87.90 | 87.90 | 70.04 | 53.58 | 50.05 |
| Cr6+ | 18.50 | 20.36 | 47.5 | 63.75 | 63.75 | 52.57 | 38.35 | 19.49 | |
| Zn2+ | 14.92 | 29.08 | 67.8 | 75.15 | 84.15 | 68.31 | 40.01 | 32.28 | |
| Nocardia sp. MORSY2014 (live cells) | Ni2+ | 5.08 | 18.40 | 49.32 | 59.65 | 61.46 | 42.70 | 38.24 | 22.70 |
| Cr6+ | 11.54 | 22.00 | 38.47 | 60.85 | 62.52 | 60.38 | 31.90 | 12.04 | |
| Zn2+ | 18.72 | 35.09 | 50.32 | 64.30 | 67.91 | 54.02 | 40.85 | 26.16 | |
| Nocardia sp. MORSY2014 (dead cells) | Ni2+ | 14.85 | 47.95 | 80.30 | 88.71 | 93.53 | 80.81 | 51.93 | 30.78 |
| Cr6+ | 20.04 | 34.78 | 75.52 | 84.57 | 95.22 | 77.10 | 70.07 | 42.05 | |
| Zn2+ | 25.37 | 50.03 | 76.87 | 87.95 | 90.37 | 85.20 | 61.44 | 50.00 | |
The effect of biosorbent dosage (0.05%–0.5%) on sorption efficiency in aqueous solutions under optimized parameters is presented in Table 6. The results indicate that when a biosorbent dosage (as in the dead biomass) was increased from 0.05% to 0.3%, the removal of Ni2+, Cr6+ and Zn2+ by both Nocardiopsis sp. MORSY1948 as well as Nocardia sp. MORSY2014 increased rapidly from 87.90%, 63.75% and 84.15% to 93.53%, 89.22% and 90.37%, respectively. Moreover, at higher concentrations of sorbent dosage (0.4% and 0.5%), it was observed that the heavy metals under study were removed from the aqueous solutions in entirety. This is because while the concentration of metal ions remains the same in solution, there are more biosorbent binding sites available at higher dosages than there are at a lower dosage, which leads to binding of all available metal ions. A similar trend with respect to the biomass effect was reported for the biosorption of Cd2+, Cu2+ and Zn2+ by S. lunalinharesii and Cr3+ by Streptomyces sp. (MB2) and S. noursei.43
Effect of biosorbet dosage (%) on biosorption efficiency (%) of the heavy metals Ni2+, Cr6+ and Zn2+ by the dead biomass of Nocardiopsis sp. MORSY1948 and Nocardia sp. MORSY2014.
| Biosorbent | Heavy metal | Biosorption efficiency (%) at different biosorbent dosage (%) | |||||
|---|---|---|---|---|---|---|---|
| 0.05 | 0.1 | 0.2 | 0.3 | 0.4 | 0.5 | ||
| Nocardiopsis sp. MORSY1948 (dead cells) | Ni2+ | 13.77 | 41.90 | 60.19 | 87.90 | 100 | 100 |
| Cr6+ | 9.52 | 23.37 | 54.48 | 63.75 | 100 | 100 | |
| Zn2+ | 16.05 | 41.52 | 72.57 | 84.15 | 100 | 100 | |
| Nocardia sp. MORSY2014 (dead cells) | Ni2+ | 15.80 | 53.20 | 67.50 | 93.53 | 100 | 100 |
| Cr6+ | 19.27 | 34.81 | 70.29 | 89.22 | 100 | 100 | |
| Zn2+ | 11.91 | 40.32 | 66.12 | 90.37 | 100 | 100 | |
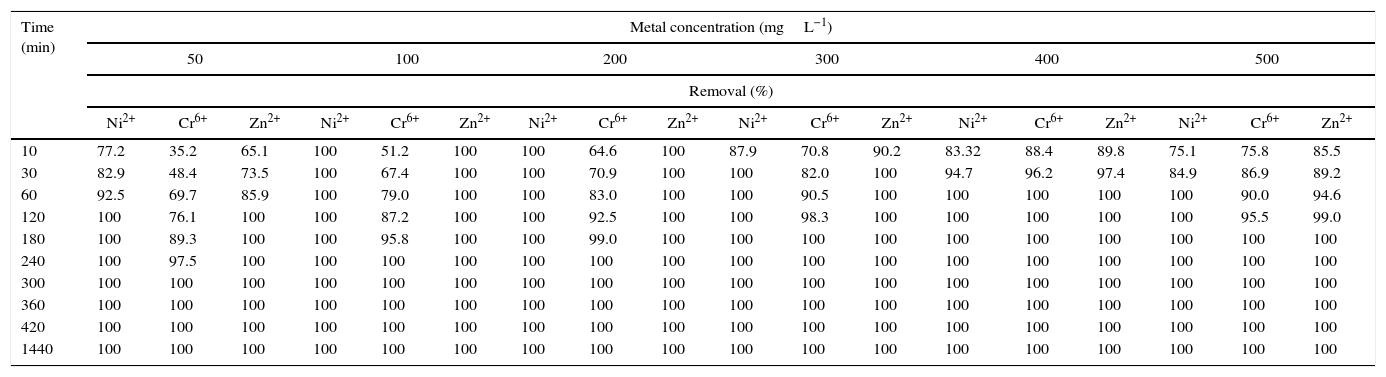
Initial concentration of heavy metal ions in the solution plays a key role as a driving force for overcoming the mass transfer resistance between the aqueous and solid phases. Equilibrium stage for both Ni2+ and Zn2+ by the biomass of strain Nocardiopsis sp. MORSY1948 was reached faster when their concentrations were increased to 100 and 200mgL−1 within 10min, respectively. For Cr6+ it took 240min for the biomass to reach equilibrium at the same concentrations (Table 7a). In case of higher metal ion concentrations (300, 400 and 500mgL−1), equilibrium point of the adsorption process was reached upon varying the contact times as follows: Ni2+ – 0, 60 and 60min; Cr6+ – 180, 60 and 180min and Zn2+ – 30, 60 and 180min, respectively. On the other hand, the time profile of the metal biosorption at different metal concentrations by the biomass of Nocardia sp. MORSY2014 strain was single, smooth and continuous leading to saturation as illustrated in Table 7b. The heavy metal ion removal efficiency reached in 10min at specific metal concentrations is as follows: 43.51%, 74.3% and 49.6% at concentration of 50mgL−1; 66.7%, 100% and 51.9% at a concentration of 100mgL−1; 75.8%, 100% and 63.75% at a concentration of 200mgL−1; 83.6%, 100% and 71.4% at a concentration of 300mgL−1; 80.7%, 85.0% and 55.3% at a concentration of 400mgL−1 and 71.0%, 81.3% and 52.0% at a concentration of 500mgL−1 for Ni2+, Cr6+, and Zn2+, respectively (Table 7b). For Nocardia sp. MORSY2014, the equilibrium stage was reached in (300, 60 and 120min), (240, 10 and 120min), (120, 10 and 120min) and (120, 10 and 60min) for Ni2+, Cr6+ and Zn2+ at concentrations equivalent to 50, 100, 200 and 300mgL−1 respectively; but at higher concentrations, i.e., 400 and 500mgL−1, it was observed that time taken to reach equilibrium was 300, 120 and 180min (Ni2+, Cr6+ and Zn2+, respectively). The increase in the bioremoval efficiency (%) observed as a result of increase in the initial concentration of Ni2+, Cr6+ and Zn2+ (mgL−1) can be attributed to an effect of an increase in the number of ions sorbed per unit strain weight. Increase in the metal ion concentration in the solution results in an increase in the diffusion of each ion at the boundary layer but post equilibrium, a reduction in the number of biomass binding sites along with slow diffusion of heavy metals to the biomass surface due to inter particle interactions and coverage of active sites results in a decrease in the removal efficiency of heavy metal ions.
Effect of different initial concentrations of metal ions on its removal by the dead biomass of Nocardiopsis sp. MORSY1948 at different contact times.
| Time (min) | Metal concentration (mgL−1) | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 50 | 100 | 200 | 300 | 400 | 500 | |||||||||||||
| Removal (%) | ||||||||||||||||||
| Ni2+ | Cr6+ | Zn2+ | Ni2+ | Cr6+ | Zn2+ | Ni2+ | Cr6+ | Zn2+ | Ni2+ | Cr6+ | Zn2+ | Ni2+ | Cr6+ | Zn2+ | Ni2+ | Cr6+ | Zn2+ | |
| 10 | 77.2 | 35.2 | 65.1 | 100 | 51.2 | 100 | 100 | 64.6 | 100 | 87.9 | 70.8 | 90.2 | 83.32 | 88.4 | 89.8 | 75.1 | 75.8 | 85.5 |
| 30 | 82.9 | 48.4 | 73.5 | 100 | 67.4 | 100 | 100 | 70.9 | 100 | 100 | 82.0 | 100 | 94.7 | 96.2 | 97.4 | 84.9 | 86.9 | 89.2 |
| 60 | 92.5 | 69.7 | 85.9 | 100 | 79.0 | 100 | 100 | 83.0 | 100 | 100 | 90.5 | 100 | 100 | 100 | 100 | 100 | 90.0 | 94.6 |
| 120 | 100 | 76.1 | 100 | 100 | 87.2 | 100 | 100 | 92.5 | 100 | 100 | 98.3 | 100 | 100 | 100 | 100 | 100 | 95.5 | 99.0 |
| 180 | 100 | 89.3 | 100 | 100 | 95.8 | 100 | 100 | 99.0 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| 240 | 100 | 97.5 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| 300 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| 360 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| 420 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| 1440 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
Effect of different initial concentrations of metal ions on its removal by the dead biomass of Nocardia sp. MORSY2014 at different contact times.
| Time (min) | Metal concentration (mgL−1) | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 50 | 100 | 200 | 300 | 400 | 500 | |||||||||||||
| Removal (%) | ||||||||||||||||||
| Ni2+ | Cr6+ | Zn2+ | Ni2+ | Cr6+ | Zn2+ | Ni2+ | Cr6+ | Zn2+ | Ni2+ | Cr6+ | Zn2+ | Ni2+ | Cr6+ | Zn2+ | Ni2+ | Cr6+ | Zn2+ | |
| 10 | 43.51 | 74.3 | 49.6 | 66.7 | 100 | 51.9 | 75.8 | 100 | 63.75 | 83.6 | 100 | 71.4 | 80.7 | 85.0 | 55.3 | 71.0 | 81.3 | 52.0 |
| 30 | 60.47 | 89.5 | 68.3 | 74.4 | 100 | 55.0 | 83.1 | 100 | 76.3 | 93.5 | 100 | 74.0 | 87.3 | 92.8 | 70.1 | 79.5 | 88.5 | 66.8 |
| 60 | 76.8 | 100 | 83.5 | 88.2 | 100 | 70.3 | 92.4 | 100 | 91.27 | 96.5 | 100 | 100 | 90.2 | 99.0 | 84.0 | 83.0 | 97.4 | 80.5 |
| 120 | 82.1 | 100 | 100 | 93.1 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 94.1 | 100 | 97.3 | 89.2 | 100 | 94.3 |
| 180 | 95.5 | 100 | 100 | 98.9 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 96.8 | 100 | 100 | 92.6 | 100 | 100 |
| 240 | 99.3 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 97.9 | 100 | 100 | 94.9 | 100 | 100 |
| 300 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| 360 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| 420 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| 1440 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
In order to assess the applicability of the optimized procedure, we attempted to apply our pilot biosorption experimental results to real wastewater samples so as to move from the experimental setup to the real-world application stage. The experiment was conducted on 100mL of pre-filtered real wastewater samples, instead of the pre-made metal solutions at different pH values (pH 2–11) under the same optimized conditions and the bioremoval data for both sets of experiments was found to be similar (Fig. 3). The maximum ion removal capacity of the dead biomass of Nocardiopsis sp. MORSY1948 for real industrial wastewater was 100% for Ni2+ and Cr6+ at pH 7.0 and the same for Zn2+ at pH 8.0. Along similar lines, the dead biomass of Nocardia sp. MORSY2014 was also observed to remove 100% of all heavy metals under study from real industrial wastewater under optimized conditions (Fig. 3). A study by Al Turk and Kiki4 recommended that halophilic Actinomycetes biomass has the potential for removal of heavy metals from raw industrial wastewater. Moreover, in a study by Chatterjee and Chandra,44 the efficiency of Geobacillus thermodenitrificans biomass in removing heavy metals from real-world effluent was determined.
Desorption Ni 2+, Cr6+ and Zn2+ heavy metals from real industrial wastewater by the dead biomass of Nocardiopsis sp. MORSY1948 and Nocardia sp. MORSY2014.")
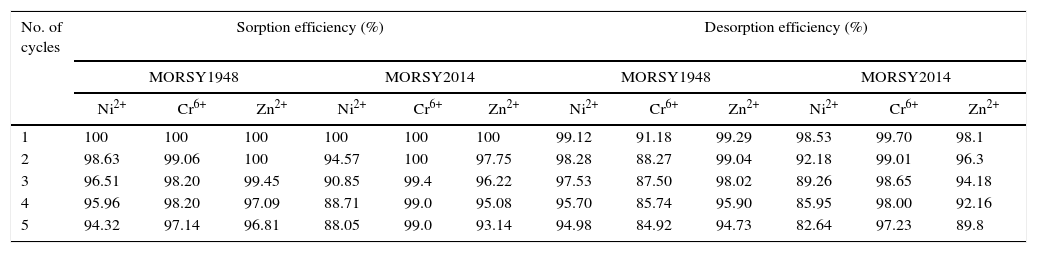
To recover heavy metals for reuse of the biosorbent, desorption efficiency should also be considered. The data presented in Table 8 illustrates that metal ions sorbed or metal ion desorbed decreased from cycle 1 to cycle 5. Biosorption efficiency was observed to reduce from 100% for Ni2+, Cr6+ and Zn2+ after the first cycle to 94.32%, 97.14% and 96.81%, respectively, after the fifth cycle. The desorption efficiency of the biomass of Nocardiopsis sp. MORSY1948 was observed to decrease from 99.12%, 91.18% and 99.29% after the first cycle to 94.98%, 84.92% and 94.73% after the fifth cycle (Table 8). The biosorption of these heavy metals by Nocardia sp. MORSY2014 was determined to be 88.05%, 99.0% and 93.14% while desorption amounts of the same metals was observed to be equal to 82.64%, 97.23% and 89.8%, respectively, after the fifth cycle (Table 8). Strong sorbent-sorbate affinity may be the main cause of differences between amount of sorbed and desorbed ions.
Desorption of Ni2+, Cr6+and Zn2+ from the biomass of Nocardiopsis sp. MORSY1948 and Nocardia sp. MORSY2014.
| No. of cycles | Sorption efficiency (%) | Desorption efficiency (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MORSY1948 | MORSY2014 | MORSY1948 | MORSY2014 | |||||||||
| Ni2+ | Cr6+ | Zn2+ | Ni2+ | Cr6+ | Zn2+ | Ni2+ | Cr6+ | Zn2+ | Ni2+ | Cr6+ | Zn2+ | |
| 1 | 100 | 100 | 100 | 100 | 100 | 100 | 99.12 | 91.18 | 99.29 | 98.53 | 99.70 | 98.1 |
| 2 | 98.63 | 99.06 | 100 | 94.57 | 100 | 97.75 | 98.28 | 88.27 | 99.04 | 92.18 | 99.01 | 96.3 |
| 3 | 96.51 | 98.20 | 99.45 | 90.85 | 99.4 | 96.22 | 97.53 | 87.50 | 98.02 | 89.26 | 98.65 | 94.18 |
| 4 | 95.96 | 98.20 | 97.09 | 88.71 | 99.0 | 95.08 | 95.70 | 85.74 | 95.90 | 85.95 | 98.00 | 92.16 |
| 5 | 94.32 | 97.14 | 96.81 | 88.05 | 99.0 | 93.14 | 94.98 | 84.92 | 94.73 | 82.64 | 97.23 | 89.8 |
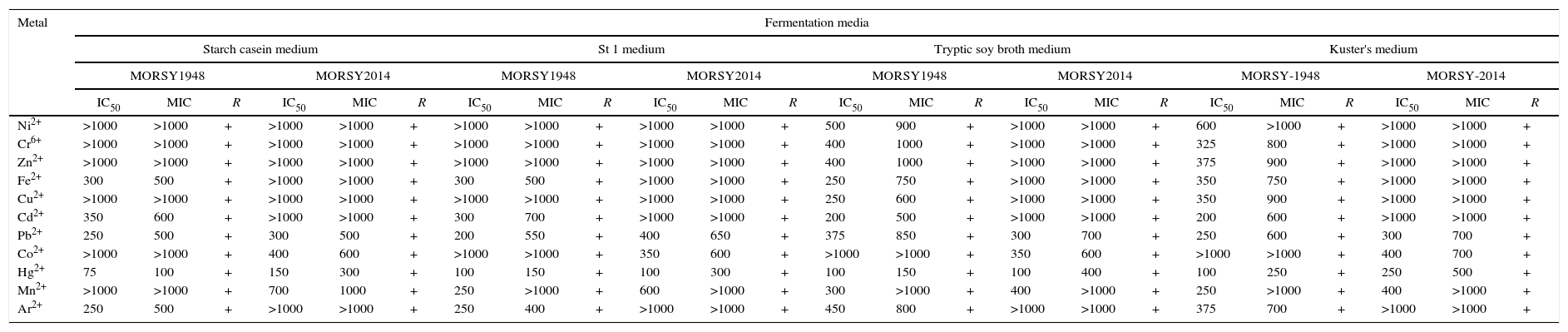
To obtain accurate comparisons, experiments to determine metal toxicity (IC50 and MIC in μgmL−1) as well as the regeneration potential of Actinomycetes were conducted using different growth media (starch casein, St 1, Kuster's and tryptic soy broth media). However, no significant differences were found in the results obtained (Table 9). The results obtained clearly revealed that in the selected biosorbent genera under study, which are representative of different taxonomic cluster groups, both Nocardiopsis sp. MORSY1948 and Nocardia sp. MORSY2014 exhibited a robust tolerance to various heavy metals that are likely to be present in contaminated sites and real industrial wastewater (Table 9). These effects can be divided into three groups. The first group wherein much greater amounts of the heavy metal ion (>1000) were required to inhibit the growth consisted of Ni2+, Cr6+, Zn2+, Cu2+, Co2+ and Mn2+ for Nocardiopsis sp. MORSY1948 and Ni2+, Cr6+, Zn2+, Fe3+, Cu2+, Cd2+ and Ar2+ for the Nocardia sp. MORSY2014. As a result of this finding, which reiterates their capacity to survive at high concentrations of Ni2+, Cr6+, Zn2+, Cu2+, Co2+, Mn2+, Fe3+, Cd2+ and Ar2+, reinforces their potential as candidates of interest for wastewater bioremediation processes especially with reference to heavy metal contaminated water bodies (Table 9). The second group of ions that had similar effects on inhibiting the yield of biomass comprised of Fe2+, Cd2+, Pb2+ and Ar2+ for Nocardiopsis sp. MORSY1948 and Pb2+, Co2+ and Hg2+ for Nocardia sp. MORSY2014 (Table 9). The third group comprising of the most toxic metal for both strains was mercury. The computed with IC50 values (75, 100, 100 and 100μgmL−1) and MIC values (100, 150, 150 and 250μgmL−1) for Nocardiopsis sp. MORSY1948 and IC50 values (150, 100, 100 and 250μgmL−1) and MIC values (300, 300, 400 and 500μgmL−1) for Nocardia sp. MORSY2014 on starch casein, St 1; tryptic soy broth and Kuster's media, respectively are mentioned in Table 9. Our data supported the theory that Actinomycete genera derived from rich heavy metal contaminated areas can be potent multi-metal resistant microorganisms. Their resistance that can be to continuous exposure to heavy metals present in wastewater results in the development of multi-metal resistance as has been reported previously.9 Some Streptomyces species such as Streptomyces strain CG252 exhibit multiple heavy metal tolerance with MIC values equal to 500μgmL−1 against Cr6+ and 1000μgmL−1 against Cu2+ along with no sensitivity to Zn2+45; similar resistance has been established in case of Streptomyces sp. VITDDK3 for arsenate, zinc, copper, mercury, cobalt, nickel and chromate46 for marine Streptomyces sp. VITSVK5 to chromium (VI and III) concentration as high as 1000mgL−1 and arsenic and lead up to 200mgL−1.8
Evaluation of IC50 (μgmL−1) and MIC (μgmL−1) of different heavy metals toward Nocardiopsis sp. MORSY1948 and Nocardia sp. MORSY2014 during fermentation course.
| Metal | Fermentation media | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Starch casein medium | St 1 medium | Tryptic soy broth medium | Kuster's medium | |||||||||||||||||||||
| MORSY1948 | MORSY2014 | MORSY1948 | MORSY2014 | MORSY1948 | MORSY2014 | MORSY-1948 | MORSY-2014 | |||||||||||||||||
| IC50 | MIC | R | IC50 | MIC | R | IC50 | MIC | R | IC50 | MIC | R | IC50 | MIC | R | IC50 | MIC | R | IC50 | MIC | R | IC50 | MIC | R | |
| Ni2+ | >1000 | >1000 | + | >1000 | >1000 | + | >1000 | >1000 | + | >1000 | >1000 | + | 500 | 900 | + | >1000 | >1000 | + | 600 | >1000 | + | >1000 | >1000 | + |
| Cr6+ | >1000 | >1000 | + | >1000 | >1000 | + | >1000 | >1000 | + | >1000 | >1000 | + | 400 | 1000 | + | >1000 | >1000 | + | 325 | 800 | + | >1000 | >1000 | + |
| Zn2+ | >1000 | >1000 | + | >1000 | >1000 | + | >1000 | >1000 | + | >1000 | >1000 | + | 400 | 1000 | + | >1000 | >1000 | + | 375 | 900 | + | >1000 | >1000 | + |
| Fe2+ | 300 | 500 | + | >1000 | >1000 | + | 300 | 500 | + | >1000 | >1000 | + | 250 | 750 | + | >1000 | >1000 | + | 350 | 750 | + | >1000 | >1000 | + |
| Cu2+ | >1000 | >1000 | + | >1000 | >1000 | + | >1000 | >1000 | + | >1000 | >1000 | + | 250 | 600 | + | >1000 | >1000 | + | 350 | 900 | + | >1000 | >1000 | + |
| Cd2+ | 350 | 600 | + | >1000 | >1000 | + | 300 | 700 | + | >1000 | >1000 | + | 200 | 500 | + | >1000 | >1000 | + | 200 | 600 | + | >1000 | >1000 | + |
| Pb2+ | 250 | 500 | + | 300 | 500 | + | 200 | 550 | + | 400 | 650 | + | 375 | 850 | + | 300 | 700 | + | 250 | 600 | + | 300 | 700 | + |
| Co2+ | >1000 | >1000 | + | 400 | 600 | + | >1000 | >1000 | + | 350 | 600 | + | >1000 | >1000 | + | 350 | 600 | + | >1000 | >1000 | + | 400 | 700 | + |
| Hg2+ | 75 | 100 | + | 150 | 300 | + | 100 | 150 | + | 100 | 300 | + | 100 | 150 | + | 100 | 400 | + | 100 | 250 | + | 250 | 500 | + |
| Mn2+ | >1000 | >1000 | + | 700 | 1000 | + | 250 | >1000 | + | 600 | >1000 | + | 300 | >1000 | + | 400 | >1000 | + | 250 | >1000 | + | 400 | >1000 | + |
| Ar2+ | 250 | 500 | + | >1000 | >1000 | + | 250 | 400 | + | >1000 | >1000 | + | 450 | 800 | + | >1000 | >1000 | + | 375 | 700 | + | >1000 | >1000 | + |
On the other hand, the regeneration ability data of both Nocardiopsis sp. MORSY1948 and Nocardia sp. MORSY2014 after subculturing from the heavy metal treated cultures into non-heavy metal media indicated that where toxic concentration of the heavy metal ions inhibited growth, they did not kill the cells as both strains were capable of regenerating with biomass comparable to that found in metal ion free control organisms (Table 9).
ConclusionsNickel, chromium and zinc have been recognized as toxic heavy metals and it is essential to remove them from wastewater both to decrease the amount of wastewater produced as well as to improve the quality of wastewater before it is released into the environment. Two Actinomycete strains, Nocardiopsis sp. MORSY1948 and Nocardia sp. MORSY2014, isolated from contaminated sites were identified as potent active biosorbents and, interestingly, our results conclusively proved that heavy metals reduction was more efficient with the dead biomass as compared to the live cells. Both strains can be used to remove toxic heavy metals from wastewater and this removal process was seen to approach 100% efficiency in aqueous solutions when the sorbent dosage was increased to 0.4% under the optimum conditions.
Conflicts of interestThe authors declare no conflicts of interest.