





INTRODUCCIÓN
Las células de Kupffer reciben el nombre del anatomista alemán Karl W. von Kupffer, que fue pionero en la identificación de las células sinusoidales hepáticas1. Estos macrófagos residentes en el hígado fueron el primer tipo celular no parenquimal caracterizado del sinusoide hepático, y su identificación contribuyó de forma decisiva a que Aschoff estableciera en 1924 el concepto de sistema reticuloendotelial del hígado2. Las células de Kupffer constituyen uno de los tipos celulares hepáticos, en términos de metabolismo y función, más activos. Así, estos macrófagos son los principales productores en el hígado de citocinas, factores de crecimiento y mediadores biológicamente activos. Las células de Kupffer ejercen funciones vitales para el organismo tales como la eliminación de sustancias extrañas y la regulación de la respuesta inflamatoria e inmunitaria. Las células de Kupffer desempeñan también un papel fisiopatológico destacado en el daño hepático por endotoxina o alcohol y en la patogénesis de la inflamación y fibrosis hepática. En la presente revisión se enumeran las principales características morfológicas y funcionales de la célula de Kupffer y se discute su participación en procesos fisiológicos y fisiopatológicos, prestando especial atención a las nuevas estrategias farmacológicas que tienen como diana estos macrófagos residentes en el hígado.
EL SINUSOIDE HEPÁTICO
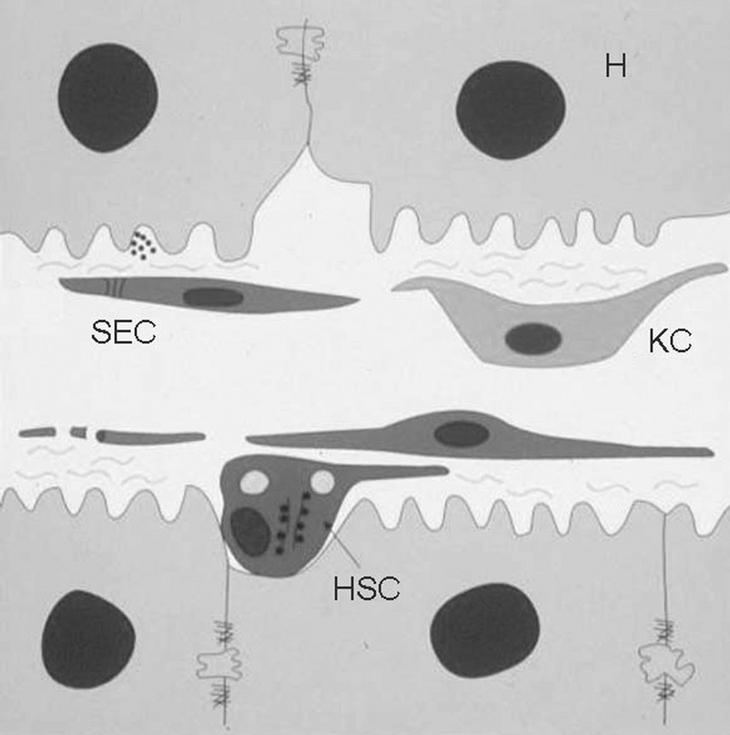
Las células de Kupffer forman parte de la unidad estructural funcional del hígado, el sinusoide hepático, que es un sistema microvascular formado por diferentes tipos celulares altamente especializados (fig. 1). Las células del sinusoide hepático se dividen en células parenquimales, los hepatocitos, que constituyen más del 80% del volumen hepático, y las células sinusoidales o no parenquimales (NPC), entre las que se encuentran las células de Kupffer; las células hepáticas estrelladas (HSC), también denominadas células de Ito, células perisinusoidales, lipocitos o células almacenadoras de grasa; las células endoteliales sinusoidales (SEC), y las células citotóxicas o pit cells.
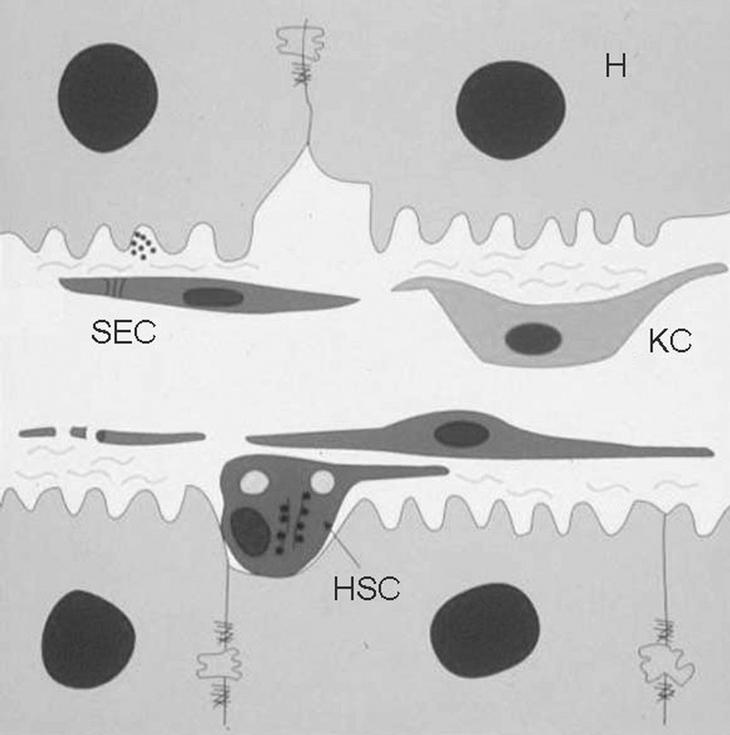
Fig. 1. Diagrama esquemático del sinusoide hepático. El sinusoide hepático está formado básicamente por 4 tipos celulares distintos. Los hepatocitos (H), que constituyen el parénquima del hígado; las células endoteliales sinusoidales (SEC), que forman el endotelio fenestrado del sinusoide; las células hepáticas estrelladas (HSC), situadas en el espacio de Disse (entre las SEC y los hepatocitos), y las células de Kupffer (KC), localizadas en el lumen del vaso.
Las células de Kupffer, que representan el 29% del total de NPC, se localizan en el lumen sinusoidal. Las células de Kupffer representan el mayor pool de macrófagos tisulares del organismo, y el hecho de que se localicen en el lumen sinusoidal las hace ser las primeras células del sistema fagocítico mononuclear que entran en contacto con el material particulado e inmunorreactivo que llega de la absorción realizada en el tracto gastrointestinal, por lo que desempeñan un importante papel en los mecanismos de defensa del organismo.
Las SEC constituyen el 48% del total de NPC y forman la pared fenestrada de los sinusoides, mientras que las HSC, que representan el 20% del total de NPC, se localizan entre los hepatocitos y las células endoteliales, en la zona conocida como espacio de Disse. Las pit cells (3% del total de NPC), linfocitos granulares de gran tamaño con actividad citotóxica y caracterizadas como las células natural killer propias del hígado, se localizan, al igual que las células de Kupffer, en el lumen sinusoidal3.
CARACTERÍSTICAS MORFOLÓGICAS DE LAS CÉLULAS DE KUPFFER
Las células de Kupffer se sitúan sobre las células endoteliales y emiten prolongaciones citoplasmáticas al espacio de Disse subendotelial, de modo que les permiten estar en contacto directo con los hepatocitos y las HSC. El estudio de la ultraestructura de estos macrófagos en el hígado de la rata muestra un contorno irregular pero de forma básicamente estrellada. La superficie celular presenta numerosos microvillis o lamelipodios e invaginaciones en forma de líneas onduladas que parecen estar implicadas en la endocitosis4. El citoplasma es rico en diferentes vesículas intracitoplasmáticas, presenta mitocondrias, un aparato de Golgi extenso, abundante retículo endoplasmático rugoso, y un conjunto de diferentes estructuras vacuolares y cuerpos densos que varían en forma, diámetro y densidad y que constituyen el desarrollado aparato lisosomal de la célula de Kupffer4. Como importantes componentes del citosqueleto de estos macrófagos encontramos un sistema muy activo de microfilamentos y microtúbulos de actina y miosina, así como filamentos de vimentina, que permiten tanto el mantenimiento de la estructura celular como el movimiento de fagocitosis y la migración celular5,6.
La identificación de las células de Kupffer se lleva a cabo habitualmente utilizando la combinación de diferentes métodos que detectan características propias del macrófago. En el hígado de rata las técnicas más utilizadas son la localización celular de la actividad peroxidasa endógena junto con la detección de la actividad fagocitaria mediante partículas de látex, oro coloidal o carbón7,8. La célula de Kupffer claramente se distingue de los monocitos circulantes por presentar la actividad peroxidasa localizada específicamente en el retículo endoplasmático y la membrana nuclear, mientras que en los monocitos su expresión sólo es a nivel lisosomal9. Sin embargo, tanto la positividad para la peroxidasa como la actividad fagocitaria son criterios que deben utilizarse con precaución por varios motivos. En primer lugar, la técnica de la peroxidasa es mucho más fiable y reproducible cuando se trata de preparaciones celulares o secciones de 1 μm de tejido incluido en resina. Además, en el hígado de rata y según el método de fijación utilizado, las SEC también pueden presentar actividad peroxidasa10. Existen también diferencias entre especies, pues en el ratón se ha observado que el 60% de las SEC presentan actividad peroxidasa11 y pueden incorporar partículas de látex12,13. En humanos, hay estudios que indican tanto la ausencia como la presencia de actividad peroxidasa en las células de Kupffer14,15.
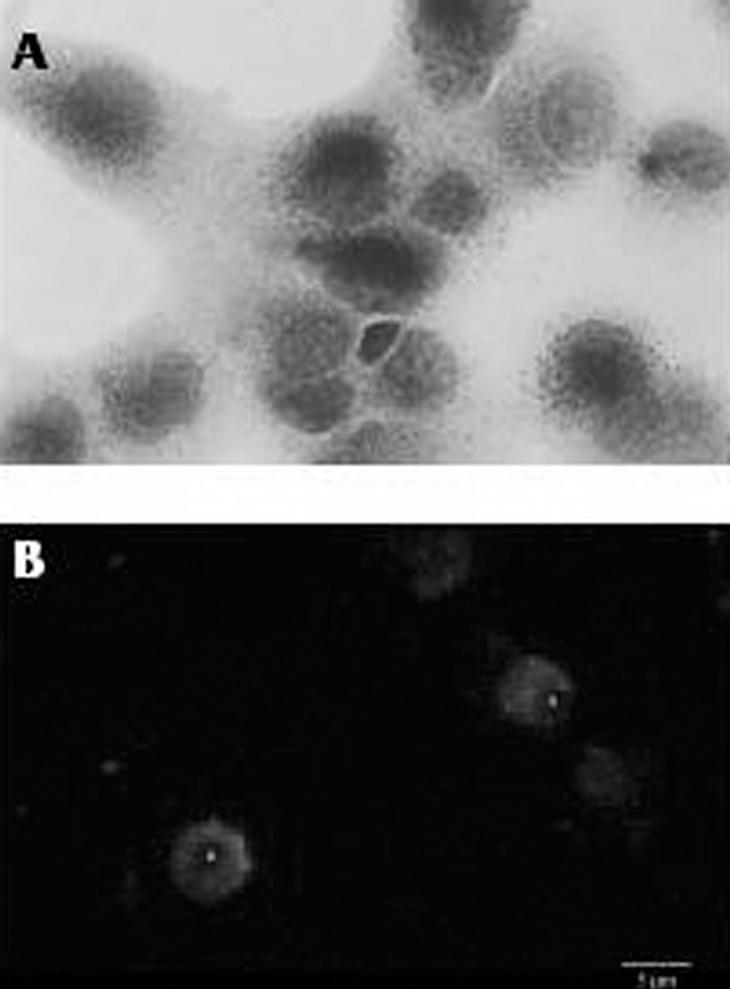
La tinción de la actividad esterasa no específica característica de los macrófagos en lugar de la peroxidasa endógena también es una técnica muy utilizada, pero necesariamente se ha de combinar con otros métodos para poder diferenciar a las células de Kupffer de macrófagos infiltrados en el hígado16. Quizá el método más fiable, pero a su vez más costoso y laborioso, es la identificación inmunohistoquímica de la células de Kupffer mediante una batería de anticuerpos monoclonales específicos para estos macrófagos. En el hígado de la rata los anticuerpos más utilizados son el ED1 y el ED2. El primero reconoce un antígeno de membrana lisosomal presente tanto en macrófagos residentes como en circulantes, y el segundo reconoce un antígeno de membrana específico de las células de Kupffer17. En nuestro laboratorio, las células de Kupffer de hígado de rata se identifican de forma rutinaria mediante una combinación del método de la esterasa no específica con la detección por métodos inmunológicos utilizando el anticuerpo ED2 (fig. 2).
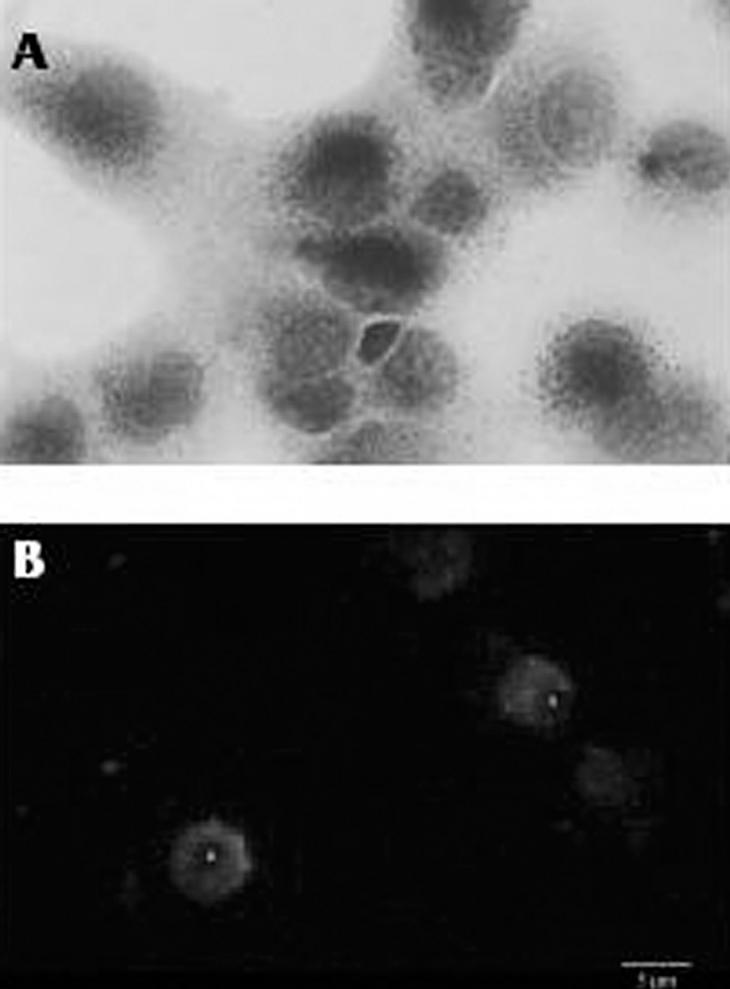
Fig. 2. Caracterización de las células de Kupffer aisladas. A) Visualización por microscopia óptica de la actividad esterasa no específica en células de Kupffer (x1.000). B) Inmunofluorescencia directa con el anticuerpo ED2 en células de Kupffer (x600).
Alternativamente, existen otros anticuerpos capaces de detectar proteínas de expresión específica en células de Kupffer en el hígado de la rata, tales como el Ki-M2R18, el UFT-419, el Ku-120 o el más recientemente caracterizado anti-KCA-321. En el ratón, los anticuerpos monoclonales F4/80 y BM8 son los más empleados22,23. Para el estudio inmunohistoquímico de las células de Kupffer en humanos se utilizan anticuerpos contra proteínas características de las células del sistema mononuclear fagocítico. Se utilizan diferentes anticuerpos policlonales y monoclonales, entre los que destacan los existentes contra la lisozima o la antitripsina α1 y los anticuerpos KP1, Mac387, EBM11 y CD6824.
CARACTERÍSTICAS FUNCIONALES DE LAS CÉLULAS DE KUPFFER
En condiciones fisiológicas, las células de Kupffer eliminan de la circulación sanguínea todo tipo de partículas extrañas, innecesarias o alteradas mediante el proceso de fagocitosis. También participan en el metabolismo de las lipoproteínas25 y desempeñan un papel clave en el proceso de captación y de detoxificación de la endotoxina que llega del flujo venoso portal26,27. Al igual que otros fagocitos mononucleares, actúan como células presentadoras de antígenos activando la respuesta inmunitaria derivada de los linfocitos T28, y también pueden desencadenar una respuesta citotóxica semejante a la realizada por las células natural killer para eliminar células tumorales circulantes29. Por último, la activación de las células de Kupffer da lugar a la secreción de potentes mediadores biológicos como radicales libres derivados del oxígeno, intermediarios del nitrógeno, numerosas citocinas y eicosanoides30. A continuación se describen con más detalle los principales aspectos funcionales de la célula de Kupffer.
Fagocitosis
La endocitosis en las células de Kupffer se puede llevar a acabo mediante 4 tipos de estructuras celulares: seudópodos fagocíticos, vacuolas, vesículas de pinocitosis e invaginaciones. La captación de partículas puede ser directa, aprovechando las particularidades físico-químicas de la sustancia, o bien mediada por receptores específicos. Las células de Kupffer expresan tanto receptores Fc como C3, de manera que se unen específicamente a los inmunocomplejos y los fagocitan31-34. También son capaces de reconocer y fagocitar glucoproteínas o glucoconjugados de oro coloidal, carbón o albúmina sérica bovina a través de un receptor de fucosa y galactosa específico de las células de Kupffer35,36. La fibronectina plasmática es una glucoproteína dimérica que también actúa como un potente agente opsonizante para facilitar la captación y la degradación de partículas extrañas por las células de Kupffer. Sin embargo, y a pesar de la existencia de lugares de unión en la membrana de las células de Kupffer37, aún no se han caracterizado receptores específicos para esta proteína en estas células. Se asume que la captación de la endotoxina está también mediada por receptor, aunque éste aún no ha sido caracterizado. La única molécula de membrana conocida que reconoce el complejo lipopolisacárido (LPS)-proteína de unión al LPS en la célula de Kupffer es el CD14, que está implicado en la transducción de señal inducida por la endotoxina38.
Metabolismo de lipoproteínas
Las lipoproteínas de baja densidad (LDL) modificadas son principalmente captadas y metabolizadas en el hígado, y las células de Kupffer, junto con las SEC, son los tipos celulares principalmente implicados39. Los receptores responsables de la captación de lipoproteínas por las células de Kupffer son los denominados receptores scavenger, también presentes de forma muy evidente en las SEC, los cuales internalizan las lipoproteínas haciéndolas susceptibles a la degradación endosomal. Las células de Kupffer y las SEC poseen la misma capacidad de captación de LDL oxidadas y acetiladas40, pero el proceso no estaría mediado por los mismos receptores. De hecho, se ha caracterizado recientemente un receptor para LDL oxidadas en las células de Kupffer homólogo a la molécula CD68 murina que no está presente en las SEC41. También se ha identificado en estos macrófagos hepáticos la expresión del receptor scavenger BI para la lipoproteína de alta densidad (HDL) rica en ésteres de colesterol, receptor que desempeña un importante papel en el llamado transporte reverso de colesterol42.
Liberación de factores de crecimiento y mediadores inflamatorios
Las células de Kupffer se activan por partículas fagocitadas y por la unión a través de receptores específicos de compuestos como ésteres de forbol, LPS, factor del complemento C5a, factores de crecimiento como GM-CSF o M-CSF, interferón γ (IFN-γ) y factor de necrosis tumoral α (TNF-α)30. La endotoxina procedente de Escherichia coli o de otras bacterias suele ser el compuesto habitualmente utilizado en el estudio de la activación de estos macrófagos38. Se considera que los signos indicativos de la activación de las células de Kupffer son el incremento de su actividad fagocítica, la mitosis, el aumento de la rugosidad de la membrana, así como la liberación de numerosos compuestos con actividad biológica. Entre éstos destacan los mediadores inorgánicos, como el ion superóxido, producidos de forma inmediata por la célula de Kupffer con el fin de inactivar y destruir el material fagocitado43. Además de los radicales libres del oxígeno, las células de Kupffer activadas sintetizan óxido nítrico, proteasas, TNF-α, interleucina (IL)-1, IL-6, IL-10, factor transformador de crecimiento beta (TGF-β), IFN-α o γ, ácido araquidónico, prostaglandinas (PG) y leucotrienos (LT) (tabla I)30. Algunos de estos productos son también estimuladores o inhibidores de su propia síntesis y la de otros mediadores. Así por ejemplo, el TNF-α estimula la síntesis de PGE2, que a su vez inhibe la liberación de TNF-α. O por ejemplo el IFN-γ, que estimula la producción de PGE2 y TNF-α, a la vez que suprime la síntesis de IL-144. También se ha comprobado tanto in vitro como in vivo que el IFN-α puede desencadenar una respuesta citotóxica de las células de Kupffer contra células tumorales45,46. Gran parte de los mediadores liberados por las células de Kupffer ejercen sus acciones de forma paracrina sobre las células adyacentes. Por ejemplo, la IL-1 por sí misma o en combinación con la IL-6 y el TNF-α no sólo modula la síntesis de ADN y proteína en los hepatocitos, sino también el metabolismo de glúcidos y lípidos y la síntesis de albúmina en estas células3. Asimismo se ha demostrado que eicosanoides como los cisteinil-leucotrienos actúan de forma paracrina sobre las HSC e inducen un incremento de la concentración intracelular de calcio y su contracción47, mientras que el LTB4 y la PGE2 liberados por la célula de Kupffer inducen la secreción de IL-8 en los hepatocitos circundantes48.

HETEROGENEIDAD MORFOLÓGICA Y FUNCIONAL DE LAS CÉLULAS DE KUPFFER
La población de células de Kupffer no se encuentra distribuida de forma homogénea a lo largo del ácino hepático, y de hecho, existe el doble de células de Kupffer en el área periportal que en el área perivenular49. Esta localización tan dependiente del grado de aporte sanguíneo hace que las células de Kupffer de la región periportal sean más grandes, posean mayor actividad lisosomal y fagocítica y generen menor cantidad de anión superóxido que las células de la región centrolobular49-51. El aislamiento de este tipo de macrófagos del hígado de rata también demuestra la existencia de subpoblaciones celulares que difieren en tamaño.
A pesar de que todas estas células presentan actividad peroxidasa endógena según el patrón característico de los macrófagos residentes, y positividad para marcadores como la esterasa no específica y ED2, la intensidad de la tinción se muestra heterogénea16. La intensidad del marcaje disminuye proporcionalmente al tamaño celular, lo que sugiere que las células más pequeñas poseen también un fenotipo más inmaduro.
La heterogeneidad fenotípica en las células de Kupffer humanas también se ha demostrado histológicamente utilizando anticuerpos monoclonales contra antígenos de macrófagos como el CD68 y el 25-F952,53. Mientras que la mayoría de los macrófagos son positivos para CD68, son menos numerosos los macrófagos maduros que presentan positividad para el 25-F9, un marcador específico de macrófagos diferenciados. Además, se encuentran tanto células doble positivas para los dos antígenos como células positivas sólo para CD6852,53. Por último, el análisis cuantitativo sugiere que la maduración de los macrófagos hepáticos es también heterogénea.
La heterogeneidad morfológica de las células de Kupffer se asocia a heterogeneidad funcional. Así, los macrófagos de mayor tamaño poseen mayor capacidad fagocítica, producen más TNF-α, IL-1 y PGE2 y poseen mayor capacidad proliferativa frente a un estímulo como el CSF-1 o el GM-CSF54. Por el contrario, los macrófagos de menor tamaño son más susceptibles a ser activados, expresan mayores cantidades de antígeno Ia, liberan mayores cantidades de óxido nítrico y anión superóxido y presentan mayor actividad citotóxica contra células tumorales55. El conjunto de estas observaciones sugiere que existe una relación estrecha entre la funcionalidad, la maduración y el tamaño de las células de Kupffer. Además, la localización en la zona periportal de los macrófagos de mayor tamaño, los que poseen más actividad fagocítica y menos actividad inflamatoria, explicaría la relativa tolerancia del hígado para los agentes inmunógenos que tienen su acceso a este órgano por la vena porta.
ORIGEN Y CINÉTICA DE LAS CÉLULAS DE KUPFFER
En la actualidad todavía existe una gran controversia sobre el origen y la capacidad proliferativa de las células de Kupffer en condiciones normales. De acuerdo con el concepto de sistema mononuclear fagocítico, los macrófagos residentes en un tejido proceden del reclutamiento de monocitos circulantes y por lo tanto serían células muy especializadas de vida corta, terminalmente diferenciadas y sin capacidad proliferativa56. Sin embargo, en estudios experimentales se ha observado que la población de macrófagos tisulares o de sus precursores no se ve afectada por la inducción de monocitopenia grave mediante la administración de Sr89 o la irradiación fraccionada de la médula ósea57-59. Estas observaciones serían compatibles con el concepto actualmente vigente de que, en condiciones normales, la población de células de Kupffer es capaz de proliferar localmente y mantenerse durante largos períodos sin el aporte de monocitos circulantes57,59,60. De hecho, estudios de ontogenia hepática demuestran la existencia en el hígado de 2 tipos de macrófagos de distinto origen hematopoyético61. Según estos estudios, existiría una población de macrófagos procedentes de los monocitos circulantes originados en la médula ósea y otra población de macrófagos tisulares resultantes de la hematopoyesis temprana acontecida durante el desarrollo embrionario61.
La capacidad proliferativa de las células de Kupffer ha sido también analizada en distintos modelos experimentales. Los resultados de estos estudios confirman el potencial mitótico de estas células en condiciones normales50,57,60,62, y la posibilidad de modular tanto in vivo como in vitro su maduración y capacidad proliferativa mediante factores solubles, como por ejemplo el M-CSF54,63,64.
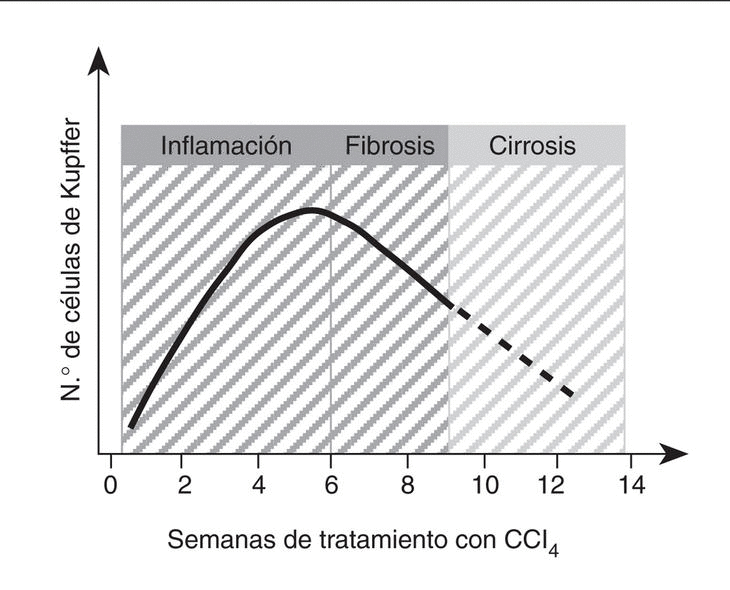
La capacidad proliferativa de las células de Kupffer adquiere mayor relevancia en el curso de la enfermedad hepática. En este sentido, se ha descrito que la población de células de Kupffer se incrementa significativamente durante las etapas iniciales de la lesión hepática65. En el caso concreto de la fibrogénesis inducida por CCl4, la población de células de Kupffer triplica su número original aproximadamente entre las 6 y las 8 semanas de tratamiento65 (fig. 3). Además, en este modelo experimental existe una correlación muy estrecha entre el número de células de Kupffer y el grado de desarrollo de fibrosis65-68. Sin embargo, una vez la cirrosis se ha establecido, el número de células de Kupffer disminuye significativamente coincidiendo con la aparición de una alteración en la función reticuloendotelial y fagocítica del hígado69. En el momento actual no se conoce con exactitud si el aumento de la población hepática de células de Kupffer durante la lesión aguda y crónica por CCl4 es consecuencia del reclutamiento de monocitos circulantes o de la proliferación local de células de Kupffer, pero posiblemente sea el resultado de ambos procesos70-72.
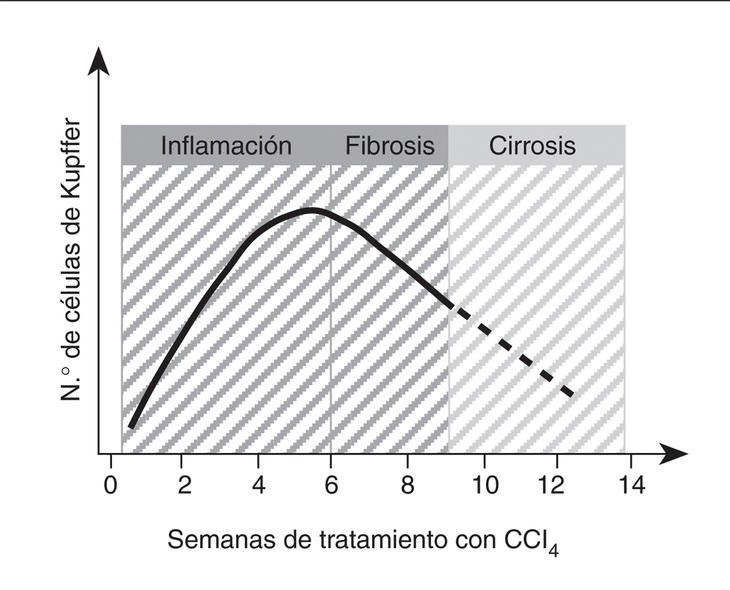
Fig. 3. Cinética de la población de células de Kupffer en el daño hepático por CCl4. Este gráfico muestra de forma resumida los datos disponibles sobre el crecimiento de la población de células de Kupffer durante el curso de la enfermedad hepática inducida por CCl4 en ratas65-68. (Véase el texto para más detalles.)
PAPEL DE LAS CÉLULAS DE KUPFFER EN LA LESIÓN HEPÁTICA
Actualmente se dispone de evidencias sólidas que demuestran la participación de las células de Kupffer en la patogénesis de la enfermedad hepática. Se sabe que estímulos como la obesidad, el consumo crónico de alcohol, la endotoxina o compuestos procedentes de la degradación de fármacos y xenobióticos inducen la activación de las células de Kupffer. Una vez activadas, las células de Kupffer liberan cantidades masivas de citocinas (IL-1, IL-6, IL-10, TNF-α), radicales libres de oxígeno y aniones superóxido, IFN-α y γy eicosanoides (PGD2, PGE2 y leucotrienos). La liberación desproporcionada de estos mediadores, junto con la secreción de enzimas lisosomales por las células de Kupffer, favorece la aparición de inflamación y necrosis en el tejido hepático. Las células de Kupffer activadas modifican también procesos metabólicos clave en las células parenquimatosas, como por ejemplo la liberación de albúmina por los hepatocitos, lo que altera la función hepática y contribuye al desarrollo de la lesión en este órgano73.
Tal como se ha apuntado anteriormente, además de ser el principal tipo celular responsable de la liberación de productos biológicamente activos, las células de Kupffer activadas incrementan significativamente su número en el hígado. Por ejemplo, en las 6 h posteriores a la administración de CCl4 se produce un aumento de la población de células de Kupffer, que presenta un pico máximo a las 72 h72. Una prueba definitiva del papel clave de las células de Kupffer en la lesión hepática es que su inactivación y su eliminación selectiva mediante cloruro de gadolinio (GdCl3) impide que aparezcan valores séricos elevados de aspartato aminotransferasa (AST) y la muerte de las células hepáticas parenquimales tras la administración de CCl474. A continuación se describen, clasificadas por la etiología del agente inductor, las principales evidencias de la participación de las células de Kupffer en diversos escenarios de daño hepático, como por ejemplo el daño inducido por la endotoxina, los fármacos, el alcohol o la isquemia-reperfusión.
Daño hepático inducido por la endotoxina
Las células de Kupffer median el daño hepático inducido por la endotoxina procedente de la pared bacteriana de Escherichia coli durante una sepsis o shock endotóxico. En condiciones normales, las células de Kupffer presentan un estado de tolerancia a la endotoxina, pero su exposición a cantidades elevadas de esta sustancia o bien la presencia de células de Kupffer en estado parcialmente activado por otras causas (p. ej., tras la administración de Corynebacterium parvum o tras hepatectomía parcial), conduce a la liberación masiva de especies reactivas del oxígeno, citocinas (principalmente TNF-α), y da lugar a fenómenos de necrosis masiva en el hígado75.
Daño hepático inducido por fármacos y xenobióticos
Junto con el riñón, la vía hepatobiliar es la principal ruta excretora de xenobióticos. Esto significa que las células hepáticas no sólo metabolizan los nutrientes procedentes del intestino y los productos metabólicos endógenos procedentes de otras partes del organismo, sino que también participan activamente en el metabolismo de los xenobióticos y especialmente de los fármacos. En condiciones normales, los fármacos son eliminados por la bilis primaria, hepatocelular o canalicular, elaborada por los hepatocitos mediante un proceso de filtración osmótica76. Sin embargo, en determinadas circunstancias, por ejemplo tras una sobredosis de fármacos, las células de Kupffer se activan y desencadenan un proceso de lesión hepática.
Un ejemplo paradigmático de daño hepático inducido por fármacos lo constituye posiblemente la intoxicación por acetaminofeno. La sobredosis accidental por acetaminofeno es una causa común de urgencia hospitalaria especialmente en el Reino Unido. La intoxicación por acetaminofeno se caracteriza por la rápida aparición de insuficiencia hepática seguida por una intensa inflamación y, finalmente, regeneración del tejido lesionado77. La hepatotoxicidad ocurre como consecuencia de la conversión por enzimas microsomales de acetaminofeno a un metabolito reactivo (la imina de N-acetil-p-benzoquinona) que se une a macromoléculas del tejido hepático e inicia una reacción necrótica78. Es durante este proceso cuando se produce una acumulación y activación de las células de Kupffer que liberan de forma masiva mediadores biológicamente activos79.
Daño hepático inducido por el alcohol
El alcohol deprime la capacidad fagocítica y la actividad lisosomal de las células de Kupffer en cultivo. Es por esta razón que clásicamente se ha considerado que las células de Kupffer no intervenían en la patogénesis del daño hepático inducido por el alcohol. Sin embargo, este concepto se ha visto modificado en los últimos años hasta el punto de que en el momento actual se considera que las células de Kupffer desempeñan un papel clave en la patogénesis de esta hepatopatía. De hecho, se ha observado un incremento del número de células de Kupffer en el hígado de pacientes con hepatitis alcohólica80. El aumento en la población de células de Kupffer se ha observado también en modelos experimentales de hepatitis alcohólica81. En estos modelos experimentales el tratamiento con GdCl3, y por tanto la inactivación de las células de Kupffer, es capaz de prevenir la elevación sérica de AST y la aparición de esteatosis, inflamación y necrosis en el tejido hepático inducida por el alcohol82.
El mecanismo por el cual las células de Kupffer participan en la lesión alcohólica no está totalmente establecido, pero se ha sugerido que puede estar relacionado con la incapacidad de estos macrófagos de eliminar la endotoxina endógena83. También se ha apuntado que las células de Kupffer se activarían por factores liberados por los hepatocitos en respuesta al alcohol. Por ejemplo, se ha descrito que las células de Kupffer en presencia de medio condicionado procedente de cultivos de hepatocitos tratados con alcohol, secretan IL-8, la cual es un potente agente quimiotáctico inductor de la infiltración de neutrófilos84. Por el contrario, otros autores sugieren la liberación de IL-8 por los hepatocitos en respuesta a factores derivados de las células de Kupffer activadas por el etanol, principalmente de IL-1 y TNF-α85,86. Por último, existen evidencias de que el daño hepático inducido por la ingesta desproporcionada de alcohol es consecuencia de la liberación de especies reactivas del oxígeno por las células de Kupffer87. También se ha descrito que la liberación de PGE2 por la célula de Kupffer en respuesta al alcohol induce la síntesis de triglicéridos por los hepatocitos, cuya acumulación contribuiría a la aparición de esteatosis88.
Daño hepático por isquemia-reperfusión
Existen evidencias morfológicas de la activación de las células de Kupffer de 1 a 3 h después de la reperfusión del hígado89. Como consecuencia de la activación de las células de Kupffer, se produce el reclutamiento de neutrófilos y la liberación por ambos tipos celulares de superóxido, TNF-α, factor activador de las plaquetas (PDGF), LTB4, PGD2, PGE2 y tromboxano B2, factores que desencadenarían la lesión tisular90,91. La implicación de las células de Kupffer en la lesión por isquemia-reperfusión se ilustra mediante la inactivación de estas células por el GdCl3, el cual es capaz de reducir la lesión hepática en modelos experimentales de reperfusión y trasplante90,91.
CÉLULAS DE KUPFFER Y FIBROGENIA
La fibrosis hepática es el resultado final de un proceso inflamatorio y de reparación del tejido que se produce en respuesta a una agresión externa del hígado. De forma muy resumida, podemos decir que la línea de acontecimientos que desencadenan la fibrosis hepática se inicia con la aparición de una respuesta inflamatoria. La inflamación es la respuesta defensiva de los organismos vivos a la agresión de agentes externos, y comprende un amplio abanico de eventos como la activación enzimática, la liberación de mediadores solubles, la extravasación de fluidos, la adhesión y migración celular y la destrucción del tejido lesionado y su posterior reparación. En condiciones normales, mediante la respuesta inflamatoria se consigue eliminar el agente causante del daño hepático. Sin embargo, en determinadas circunstancias, especialmente si el daño se produce de forma crónica y continuada, la respuesta inflamatoria no se resuelve de forma adecuada y se autoperpetúa en el tiempo, dando lugar a procesos de secreción inadecuada de matriz extracelular y a la aparición de fibrosis92.
El principal tipo celular responsable de la secreción de proteínas de matriz son las HSC en su estado activado. Por este motivo, la activación y proliferación de las HSC constituye un proceso clave en el desarrollo de fibrosis hepática92,93. Las células de Kupffer, mediante la liberación de citocinas, eicosanoides y factores de crecimiento, desempeñan un papel primordial en la activación y proliferación de las HSC94. Se ha demostrado que el medio condicionado procedente de cultivos de células de Kupffer promueve la proliferación de los HSC y la síntesis de colágeno, proteoglicanos y hialuronato95-98, efecto que es más acentuado si las células de Kupffer proceden de hígados de ratas tratadas con CCl496,98. Se han identificado numerosos compuestos liberados por las células de Kupffer que pueden ejercer este efecto proliferativo y estimulador de las HSC, y entre ellos destacan el TGF-β, eicosanoides, PDGF, TNF-α, IL-1 e IFN-γ. (Para una revisión exhaustiva, véase la referencia 93 de la bibliografía.)
Las células de Kupffer contribuyen también a la activación de las HSC mediante la degradación de la matriz extracelular subendotelial, la cual, como es sabido, mantiene a las HSC en su estado quiescente93. En concreto, las células de Kupffer activadas son las principales productoras de colagenasa tipo IV/gelatinasa tipo B con capacidad de digerir esta matriz extracelular, y por tanto de inducir la activación, proliferación y migración de las HSC73,99.
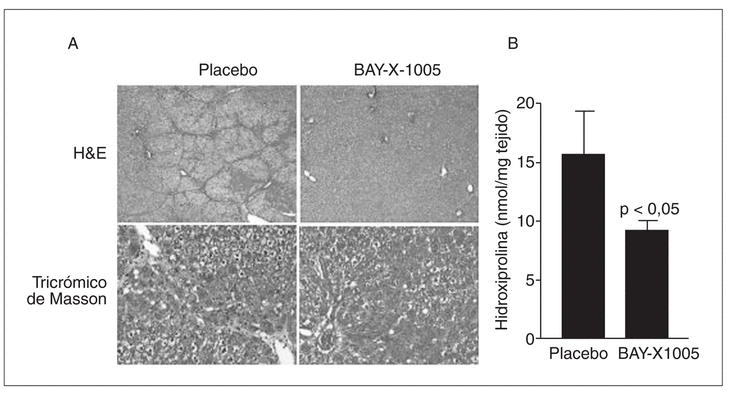
Tal como se ha comentado anteriormente, es importante señalar que durante las etapas iniciales de la fibrogenia la población de células de Kupffer se incrementa significativamente65. Una de las estrategias antifibróticas emprendidas en nuestro laboratorio contempla la posibilidad de bloquear el crecimiento de las células de Kupffer con inhibidores de la 5-lipoxigenasa (5-LO), la enzima clave en la síntesis de leucotrienos. La base racional de esta estrategia se basa principalmente en dos conceptos: el primero es que determinados tipos celulares requieren la presencia de una 5-LO metabólicamente activa para garantizar su supervivencia, y el segundo es que en el hígado esta enzima se halla exclusivamente en las células de Kupffer47. Los resultados obtenidos hasta el momento son muy esperanzadores, ya que hemos observado que la inhibición de la vía de la 5-LO induce muerte celular por apoptosis en las células de Kupffer y que la inhibición farmacológica de esta vía de metabolismo del ácido araquidónico in vivo reduce el grado de fibrosis en el modelo experimental de CCl471 (fig. 4).
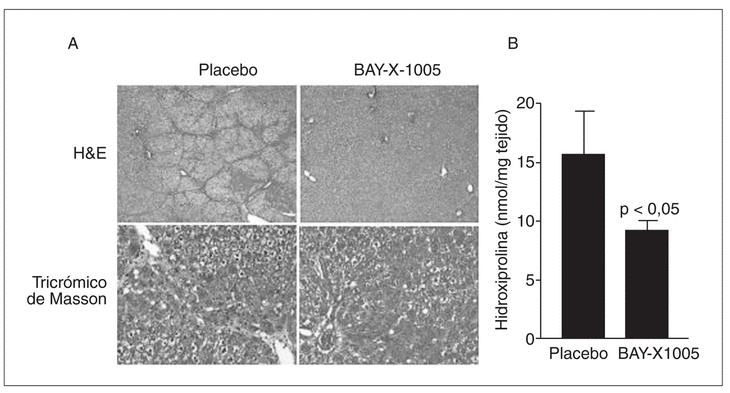
Fig. 4. Efectos antifibróticos de la inhibición de la 5-lipoxigenasa (5-LO) en ratas tratadas con CCl4. A) Secciones histológicas de tejido hepático teñidas con hematoxilina-eosina (H&E) (paneles superiores,x25) o tricrómico de Masson (paneles inferiores, x125) de animales tratados con CCl4 que recibieron durante 8 semanas una dosis diaria del inhibidor de la vía de la 5-LO BAY-X-1005 (100 mg/kg) o placebo (carboximetilcelulosa). B) Contenido hepático en hidroxiprolina en estos mismos animales.
CÉLULAS DE KUPFFER Y RESPUESTA INMUNITARIA
La respuesta inmunitaria ocurre posteriormente a la respuesta inflamatoria, y en ella las células inmunológicamente competentes (monocitos-macrófagos y linfocitos) se activan en respuesta a sustancias liberadas durante la fase inflamatoria aguda (IL-1, IL-2, TNF-α, leucotrienos). El desarrollo de la respuesta inmunitaria suele ser beneficioso para el organismo si el agente agresor es neutralizado o fagocitado. Sin embargo, si la respuesta inmunitaria no resuelve bien este proceso, aparece la fase crónica de inflamación, que es cuando se liberan numerosos mediadores (GM-CSF, TNF-α, IFN-γ, PDGF) que autoperpetúan el proceso inflamatorio. Es por esta razón que las células de Kupffer desempeñan un papel clave en la defensa inmunitaria innata. Se considera que las células de Kupffer están también implicadas en la patogénesis del rechazo agudo del hígado, puesto que estos macrófagos son capaces de modular la respuesta inmunitaria al actuar como células presentadoras de antígenos, de regular la activación de los linfocitos T y de participar en el desarrollo de tolerancia a antígenos solubles de origen gastrointestinal100,101.
CÉLULAS DE KUPFFER Y REGENERACIÓN HEPÁTICA
En condiciones normales, el recambio celular en el hígado es un proceso constante aunque no evidente. Sin embargo, asociado a la resolución de la lesión hepática producida, por ejemplo, por un virus o un fármaco, se observa un espectacular incremento de la proliferación hepática y principalmente de los hepatocitos, que da lugar al proceso denominado de regeneración hepática. Quizá el modelo experimental más utilizado para investigar la regeneración hepática sea la hepatectomía parcial (eliminación del 70% de la masa hepática). Se ha observado que de 1 a 3 días después de practicar la hepatectomía, se produce un pico de síntesis de ADN en las células no parenquimales del hígado, incluidas las células de Kupffer102. Esta expansión de la población de células de Kupffer procede principalmente de la proliferación local de estos macrófagos103. Por tanto, y de forma paralela a la proliferación de los hepatocitos, durante el proceso de regeneración hepática se produce también la proliferación de las células de Kupffer.
La regeneración hepática es un proceso altamente regulado y sincronizado, con el fin de recuperar la masa hepática perdida. En esta regulación intervienen diversos factores de crecimiento e interacciones célula a célula, y en ella las células de Kupffer desempeñan un papel relevante. En este contexto estos macrófagos secretan factores de crecimiento tanto estimuladores como inhibidores de la síntesis de ADN y la proliferación de los hepatocitos. Así, la síntesis del mitógeno más potente para los hepatocitos, el factor de crecimiento de los hepatocitos (HGF), se produce durante el proceso de regeneración hepática en las células no parenquimales (en orden de mayor a menor grado de participación: HSC, células de Kupffer y SEC)104. Otros factores de crecimiento secretados por las células de Kupffer son el factor de crecimiento de la insulina (IGF) y el factor de unión de la heparina (HBGF). Mención especial requiere el TGF-α. Se considera que tras la hepatectomía, la secreción paracrina de TGF-α por las células de Kupffer es un factor necesario para el inicio de la proliferación de los hepatocitos105-107. Sin embargo, en etapas más tardías de la regeneración hepática, el TGF-β secretado de forma casi exclusiva en las células de Kupffer actuaría regulando negativamente la proliferación de los hepatocitos108. Aunque estos resultados parecen controvertidos, conceptualmente indican que factores liberados por las células de Kupffer modulan la proliferación de los hepatocitos. Existen evidencias que apoyan esta idea, como por ejemplo el incremento en la proliferación de los hepatocitos tras la depleción selectiva de las células de Kupffer con diclorometileno difosfonato encapsulado en liposomas109.
Nuestro laboratorio está parcialmente financiado por el Ministerio de Educación y Ciencia (SAF 03/0586) y por el Instituto de Salud Carlos III (C03/02).

