






Para aumentar la resistencia de las plantas a los patógenos se requiere de un trabajo interdisciplinario de las ciencias biológicas y agrícolas. En este escenario, la fitopatología se ha beneficiado del encuentro de disciplinas biológicas claves, como la ecología, la evolución y la biología molecular. Sin embargo, este encuentro no ha estado exento de controversia entre los académicos de estas disciplinas, quienes a menudo han otorgado diferentes significados a un mismo concepto. Este documento tiene como propósito ofrecer una visión abarcativa y moderna de la resistencia de las plantas a los patógenos, enmarcada dentro del concepto clásico de resistencia vertical y horizontal propuesto por Van der Plank. Este concepto tiene implicaciones en áreas centrales para la agricultura, como el fitomejoramiento y la genética vegetal, lo que justifica su uso como el marco conceptual indicado. Dentro de este contexto se explican otros conceptos clásicos de la fitopatología en combinación con descripciones modernas de la interacción entre plantas y patógenos, y se discute cómo estos conceptos están relacionados con la resistencia cuantitativa y de campo. Asimismo, esta revisión tiene un propósito adicional como aporte a la enseñanza de la fitopatología en la América de habla hispana, dado que en esta región existe, además de la barrera interdisciplinaria, un factor adicional y persistente que impide llevar la discusión a un terreno común: la baja competencia lingüística en el idioma inglés en todos los niveles educativos. Esta revisión pretende ser una herramienta de apoyo para los docentes de fitopatología de la región, interesados en explicar los conceptos modernos de resistencia a patógenos en plantas.
Understanding plant resistance requires an interdisciplinary effort between biological and agricultural sciences. In this setting, phytopathology has experienced an upsurge of interest from researchers and scholars in the disciplines of ecology, evolution and molecular biology. This encounter did neither avoid misunderstandings among scholars nor the use of similar concepts with different meanings. The purpose of this paper is to offer a modern comprehensive view of plant resistance against pathogens using a classical phytopathology concept as framework: Van der Plank s concept of horizontal and vertical resistance. This concept is used in other agricultural science disciplines (plant breeding and genetics), supporting why it is a proper framework for explaining plant resistance. Within this frame, other classical phytopathologycal concepts are explained in combination with modern model descriptions of plant-pathogen interactions and how all these concepts are related with quantitative and field resistance. This review is written in Spanish because it serves an additional purpose. In the Spanish-speaking America, besides interdisciplinarity, phytopathology as an academic discipline faces another challenge: the students’ low-English language proficiency. In this regard, this review intends to become a companion guide for plant-pathology teachers in the region interested in providing an insight into the modern concepts of plant resistance.
El estudio de la resistencia de las plantas a los patógenos requiere un trabajo interdisciplinario entre las ciencias biológicas y las ciencias agrícolas y, en consecuencia, entre áreas del conocimiento relativamente distintas: por un lado, la de la fitopatología, una ciencia aplicada y, por otro, la de la interacción planta-patógeno, un fenómeno biológico con implicaciones ecológicas, evolutivas y moleculares. Los conceptos de resistencia a patógenos han estado sujetos a reiteradas controversias entre los académicos de estas áreas y suelen ser de difícil comprensión para los estudiantes de pregrado y posgrado, así como para los docentes e investigadores no familiarizados con las enfermedades de las plantas. Las dificultades también persisten entre expertos porque, en múltiples ocasiones, términos similares son usados para definir conceptos distintos en interacciones que no son estrictamente patogénicas desde el punto de vista agronómico (porque no causan daño visible o disminución de rendimiento) o que representan diferentes medidas físicas2,5,9,36,48,52,53.
En la América de habla hispana, además de la barrera interdisciplinaria, existe un factor adicional persistente que impide llevar la discusión a un terreno común y lograr una comprensión conjunta del tema. Ese factor es la escasa competencia lingüística en el idioma inglés que se advierte en todos los niveles educativos, desde el básico escolar hasta el universitario, y ello impacta en el desarrollo profesional de los graduados10,12,13,22. En la experiencia del autor como docente e investigador, es recurrente observar cómo los estudiantes e incluso muchos investigadores carecen de las suficientes competencias para acceder a la información publicada en inglés. La literatura científica anglosajona contiene múltiples revisiones sobre el concepto de resistencia, pero estas no pueden ser consultadas debido a las limitaciones lingüísticas de sus potenciales lectores. En este contexto, esta revisión tiene 2propósitos: ofrecer una visión comprensible de la evolución de los conceptos de resistencia, especialmente el de resistencia cuantitativa, y servir como documento de estudio y referencia para estudiantes, docentes e investigadores de habla hispana.
Los conceptos de resistencia a patógenos involucran términos con diferencias idiomáticas que merecen ser mencionadas. Una de ellas es el valor semántico (significado) de la palabra huésped. En el español coloquial, la palabra huésped tiene 2significados: el más frecuente hace referencia a la «persona alojada en casa ajena» y el segundo, escasamente usado en la actualidad, alude a la «persona que hospeda en su casa a otra»44. En concordancia con el primer uso, en la actualidad más extendido, la palabra «huésped» equivale semánticamente al término parásito o patógeno, y consideramos que así debe ser usada en el contexto científico. Su contraparte es el hospedero o anfitrión: el organismo donde se aloja el parásito o patógeno. En consecuencia, la relación objeto de la fitopatología estaría adecuadamente descrita en español como el estudio de la interacción hospedero-patógeno. La figura 1 ilustra estos conceptos usando una enfermedad común en Sudamérica en los cultivos de algodón, la ramulosis, provocada por Colletotrichum gossypii var. cephalosporioides32.
 e hifas secundarias (en las células que rodean la célula donde se inició el proceso infectivo). C. Lesiones tipo estrella en las hojas de algodón provocadas por el patógeno. Las lesiones tipo estrella son el resultado de la necrosis intensa que sufre el punto central donde la infección comenzó. Este es el primer síntoma de ramulosis, que continúa con la colonización de los meristemas de la planta y su necrosis, lo que lleva al enanismo y la pérdida total de la cosecha.")
Interacción entre un hospedero vegetal y un patógeno. El ejemplo está basado en una enfermedad específica para América del Sur: la ramulosis del algodón, provocada por Colletotrichum gossypii var. cephalosporioides. A. Colonia y esporas de este patógeno. B. Representación esquemática de C. gossypii var. cephalosporioides invadiendo células de hojas de algodón. La imagen muestra la espora germinada, la formación del apresorio en la superficie de la planta y la formación de hifas multilobuladas (en la primera célula infectada) e hifas secundarias (en las células que rodean la célula donde se inició el proceso infectivo). C. Lesiones tipo estrella en las hojas de algodón provocadas por el patógeno. Las lesiones tipo estrella son el resultado de la necrosis intensa que sufre el punto central donde la infección comenzó. Este es el primer síntoma de ramulosis, que continúa con la colonización de los meristemas de la planta y su necrosis, lo que lleva al enanismo y la pérdida total de la cosecha.
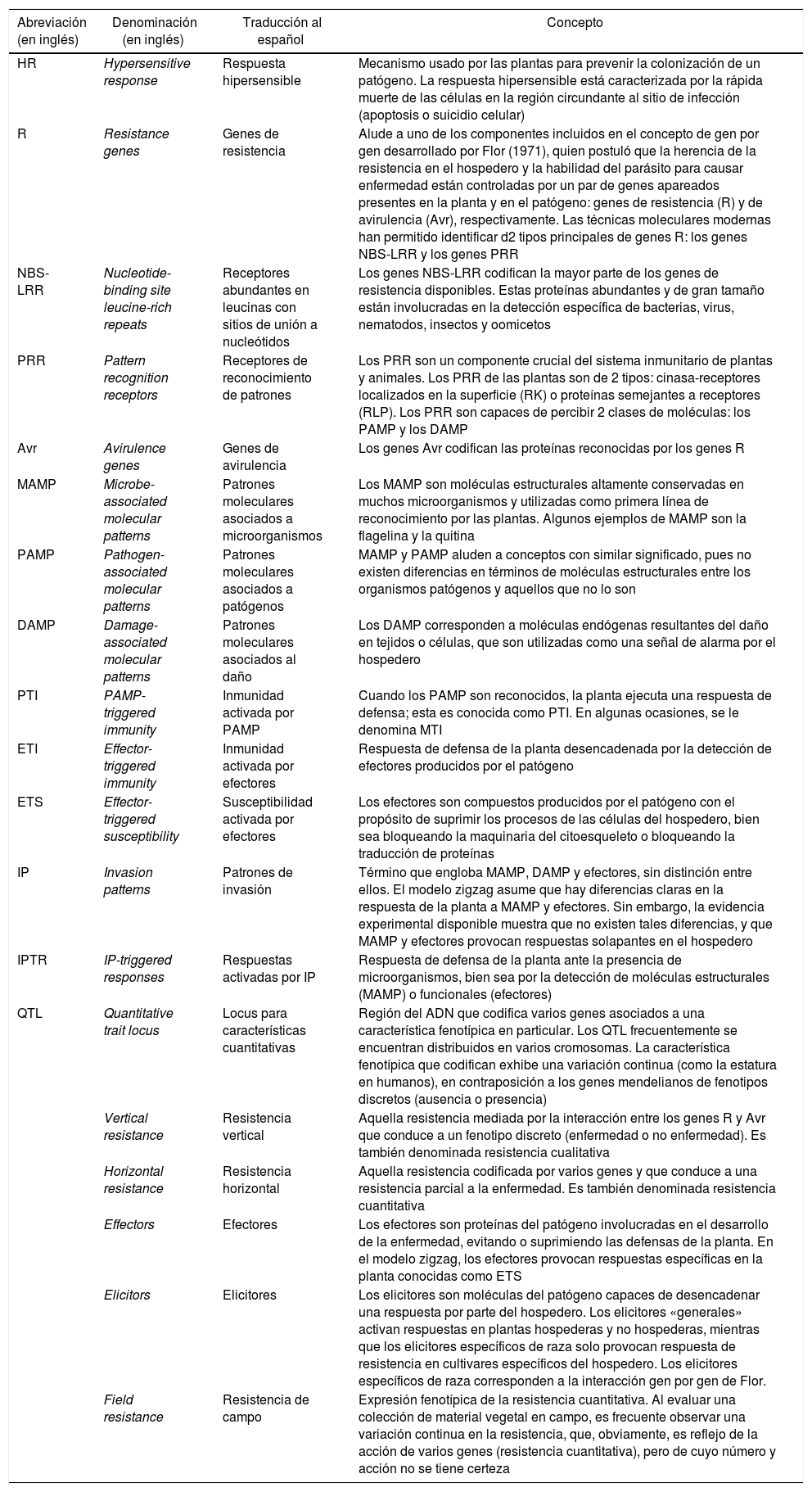
En la tabla 1 se presenta una serie de conceptos vinculados con la interacción entre las plantas y sus patógenos4,6,29,31,35,41,45, con el detalle de cuáles son las abreviaciones usadas en inglés para nombrarlos y cuál es su traducción al español, a fin de unificar criterios y facilitar la lectura en ambos idiomas.
Nómina de abreviaciones y descripción de conceptos usadas en este documento
| Abreviación (en inglés) | Denominación (en inglés) | Traducción al español | Concepto |
|---|---|---|---|
| HR | Hypersensitive response | Respuesta hipersensible | Mecanismo usado por las plantas para prevenir la colonización de un patógeno. La respuesta hipersensible está caracterizada por la rápida muerte de las células en la región circundante al sitio de infección (apoptosis o suicidio celular) |
| R | Resistance genes | Genes de resistencia | Alude a uno de los componentes incluidos en el concepto de gen por gen desarrollado por Flor (1971), quien postuló que la herencia de la resistencia en el hospedero y la habilidad del parásito para causar enfermedad están controladas por un par de genes apareados presentes en la planta y en el patógeno: genes de resistencia (R) y de avirulencia (Avr), respectivamente. Las técnicas moleculares modernas han permitido identificar d2 tipos principales de genes R: los genes NBS-LRR y los genes PRR |
| NBS-LRR | Nucleotide-binding site leucine-rich repeats | Receptores abundantes en leucinas con sitios de unión a nucleótidos | Los genes NBS-LRR codifican la mayor parte de los genes de resistencia disponibles. Estas proteínas abundantes y de gran tamaño están involucradas en la detección específica de bacterias, virus, nematodos, insectos y oomicetos |
| PRR | Pattern recognition receptors | Receptores de reconocimiento de patrones | Los PRR son un componente crucial del sistema inmunitario de plantas y animales. Los PRR de las plantas son de 2 tipos: cinasa-receptores localizados en la superficie (RK) o proteínas semejantes a receptores (RLP). Los PRR son capaces de percibir 2 clases de moléculas: los PAMP y los DAMP |
| Avr | Avirulence genes | Genes de avirulencia | Los genes Avr codifican las proteínas reconocidas por los genes R |
| MAMP | Microbe-associated molecular patterns | Patrones moleculares asociados a microorganismos | Los MAMP son moléculas estructurales altamente conservadas en muchos microorganismos y utilizadas como primera línea de reconocimiento por las plantas. Algunos ejemplos de MAMP son la flagelina y la quitina |
| PAMP | Pathogen-associated molecular patterns | Patrones moleculares asociados a patógenos | MAMP y PAMP aluden a conceptos con similar significado, pues no existen diferencias en términos de moléculas estructurales entre los organismos patógenos y aquellos que no lo son |
| DAMP | Damage-associated molecular patterns | Patrones moleculares asociados al daño | Los DAMP corresponden a moléculas endógenas resultantes del daño en tejidos o células, que son utilizadas como una señal de alarma por el hospedero |
| PTI | PAMP-triggered immunity | Inmunidad activada por PAMP | Cuando los PAMP son reconocidos, la planta ejecuta una respuesta de defensa; esta es conocida como PTI. En algunas ocasiones, se le denomina MTI |
| ETI | Effector-triggered immunity | Inmunidad activada por efectores | Respuesta de defensa de la planta desencadenada por la detección de efectores producidos por el patógeno |
| ETS | Effector-triggered susceptibility | Susceptibilidad activada por efectores | Los efectores son compuestos producidos por el patógeno con el propósito de suprimir los procesos de las células del hospedero, bien sea bloqueando la maquinaria del citoesqueleto o bloqueando la traducción de proteínas |
| IP | Invasion patterns | Patrones de invasión | Término que engloba MAMP, DAMP y efectores, sin distinción entre ellos. El modelo zigzag asume que hay diferencias claras en la respuesta de la planta a MAMP y efectores. Sin embargo, la evidencia experimental disponible muestra que no existen tales diferencias, y que MAMP y efectores provocan respuestas solapantes en el hospedero |
| IPTR | IP-triggered responses | Respuestas activadas por IP | Respuesta de defensa de la planta ante la presencia de microorganismos, bien sea por la detección de moléculas estructurales (MAMP) o funcionales (efectores) |
| QTL | Quantitative trait locus | Locus para características cuantitativas | Región del ADN que codifica varios genes asociados a una característica fenotípica en particular. Los QTL frecuentemente se encuentran distribuidos en varios cromosomas. La característica fenotípica que codifican exhibe una variación continua (como la estatura en humanos), en contraposición a los genes mendelianos de fenotipos discretos (ausencia o presencia) |
| Vertical resistance | Resistencia vertical | Aquella resistencia mediada por la interacción entre los genes R y Avr que conduce a un fenotipo discreto (enfermedad o no enfermedad). Es también denominada resistencia cualitativa | |
| Horizontal resistance | Resistencia horizontal | Aquella resistencia codificada por varios genes y que conduce a una resistencia parcial a la enfermedad. Es también denominada resistencia cuantitativa | |
| Effectors | Efectores | Los efectores son proteínas del patógeno involucradas en el desarrollo de la enfermedad, evitando o suprimiendo las defensas de la planta. En el modelo zigzag, los efectores provocan respuestas específicas en la planta conocidas como ETS | |
| Elicitors | Elicitores | Los elicitores son moléculas del patógeno capaces de desencadenar una respuesta por parte del hospedero. Los elicitores «generales» activan respuestas en plantas hospederas y no hospederas, mientras que los elicitores específicos de raza solo provocan respuesta de resistencia en cultivares específicos del hospedero. Los elicitores específicos de raza corresponden a la interacción gen por gen de Flor. | |
| Field resistance | Resistencia de campo | Expresión fenotípica de la resistencia cuantitativa. Al evaluar una colección de material vegetal en campo, es frecuente observar una variación continua en la resistencia, que, obviamente, es reflejo de la acción de varios genes (resistencia cuantitativa), pero de cuyo número y acción no se tiene certeza |
La resistencia a patógenos en fitopatología ha estado definida históricamente con los conceptos de «gen por gen» de Flor17 y «resistencia vertical y horizontal» de Van der Plank55-57. Estos conceptos constituyeron la primera aproximación teórica a la noción de resistencia a patógenos en plantas y aún son usados en la literatura moderna para describir modelos generales y casos específicos de resistencia1.
La interacción gen por gen de FlorEn los años cincuenta, Flor desarrolló uno de los conceptos clásicos de la fitopatología: la denominada hipótesis de interacción gen por gen17. Este concepto propone que para cada gen del hospedero que confiere resistencia hay un gen correspondiente que confiere patogenicidad en el parásito. En consideración del marco conceptual de esta hipótesis, es posible construir genotipos hipotéticos del hospedero y el patógeno sin desarrollar estudios genéticos, tan solo determinando las reacciones de poblaciones del patógeno a un rango de variedades del hospedero (los diferenciales de papa para Phytophthora infestans, por ejemplo).
A los genes de resistencia se les denomina R y a sus contrapartes en el patógeno, genes de avirulencia (Avr). Los genes R y los genes Avr interactúan uno a uno. Cuando un gen R identifica un gen Avr en el patógeno, activa un proceso de muerte celular en la célula infectada, la denominada respuesta hipersensible. La muerte de la célula infectada por el patógeno detiene por completo el proceso de infección y colonización, y otorga así resistencia a la planta.
El concepto de resistencia vertical y horizontal de Van der PlankEn los años sesenta, Van der Plank planteó un modelo para explicar la existencia de razas en los patógenos, la resistencia de los hospederos y las diferencias resultantes de sus interacciones55-57 Van der Plank clasificó la resistencia del hospedero en 2tipos básicos (fig. 2):
 Gota o tizón tardío de la papa. El diagrama de resistencia vertical muestra la interacción entre diferenciales de papa (Solanum tuberosum) que contienen genes R y razas fisiológicas de Phytophthora infestans. Actualmente, existen diferenciales de papa transgénicos que contienen un solo gen de resistencia y mantienen el mismo genotipo de papa60, haciendo que el esquema descrito sean una aproximación realista del fenómeno experimental observable. Para cada diferencial existe una población del patógeno que es portador del gen de avirulencia respectivo (Avr) que facilita la detección del patógeno por la planta y el desarrollo de la respuesta hipersensible. Las patógenos no portadores del gen Avr respectivo pueden infectar la planta de manera exitosa. Las razas del patógeno son designadas en función de los diferenciales que pueda infectar (y, en consecuencia, los genes Avr portados). B) Roya foliar del trigo. La resistencia vertical descrita muestra la interacción entre los diferenciales de trigo que portan los genes Lr12, Lr13 y Lr22 y las razas fisiológicas del hongo causante de la roya de la hoja (Puccinia triticina). A diferencia del ejemplo del tizón tardío, las razas son designadas en función de los diferenciales de trigo que pueden ser infectados. Hasta el momento ningún gen Avr ha sido identificado, pero los estudios genéticos apoyan una interacción entre patógeno y hospedero siguiendo el modelo gen por gen de Flor16. En la interacción descrita, la resistencia es observada como pecas necróticas con nula o escasa producción de uredinios que indican reacción hipersensible. El desarrollo de la enfermedad (hospedero susceptible es visualizado como la presencia de uredinios sin evidencia de clorosis. Además de la respuesta hipersensible, la resistencia puede observarse como no desarrollo de lesión alguna y en este caso se denomina inmunidad. En el caso de la resistencia horizontal, la representación ilustra la interacción mediada por los genes Lr34, Lr46 y Lr67. Estos genes controlan una respuesta de la planta que retarda el desarrollo del patógeno, reduciendo la velocidad de aparición de las pústulas de la roya, lo que resulta en un menor número de lesiones. La resistencia mediada por estos genes no solo es efectiva contra P. triticina sino contra otras 2 especies de patógenos causantes de roya (Pucciniastriiformis f. sp. tritici y Puccinia graminis f. sp. tritici) y contra el mildiu del trigo (Blumeria graminis f. sp. tritici). Los genes de resistencia vertical y horizontal descritos en el caso de la roya solo se expresan durante la adultez de la planta. Para la resistencia horizontal, en los 2 ejemplos usados, la población del patógeno no está dividida en razas porque no existen interacciones específicas entre el patógeno y las variedades de la planta. Las variedades del hospedero, en cambio, exhiben diferentes respuestas al patógeno que resultan en diferentes niveles de enfermedad. En el caso de la resistencia vertical, solo existen 2 opciones: ausencia o presencia de la enfermedad.")
Representación esquemática de los conceptos de resistencia vertical y horizontal. A) Gota o tizón tardío de la papa. El diagrama de resistencia vertical muestra la interacción entre diferenciales de papa (Solanum tuberosum) que contienen genes R y razas fisiológicas de Phytophthora infestans. Actualmente, existen diferenciales de papa transgénicos que contienen un solo gen de resistencia y mantienen el mismo genotipo de papa60, haciendo que el esquema descrito sean una aproximación realista del fenómeno experimental observable. Para cada diferencial existe una población del patógeno que es portador del gen de avirulencia respectivo (Avr) que facilita la detección del patógeno por la planta y el desarrollo de la respuesta hipersensible. Las patógenos no portadores del gen Avr respectivo pueden infectar la planta de manera exitosa. Las razas del patógeno son designadas en función de los diferenciales que pueda infectar (y, en consecuencia, los genes Avr portados). B) Roya foliar del trigo. La resistencia vertical descrita muestra la interacción entre los diferenciales de trigo que portan los genes Lr12, Lr13 y Lr22 y las razas fisiológicas del hongo causante de la roya de la hoja (Puccinia triticina). A diferencia del ejemplo del tizón tardío, las razas son designadas en función de los diferenciales de trigo que pueden ser infectados. Hasta el momento ningún gen Avr ha sido identificado, pero los estudios genéticos apoyan una interacción entre patógeno y hospedero siguiendo el modelo gen por gen de Flor16. En la interacción descrita, la resistencia es observada como pecas necróticas con nula o escasa producción de uredinios que indican reacción hipersensible. El desarrollo de la enfermedad (hospedero susceptible es visualizado como la presencia de uredinios sin evidencia de clorosis. Además de la respuesta hipersensible, la resistencia puede observarse como no desarrollo de lesión alguna y en este caso se denomina inmunidad. En el caso de la resistencia horizontal, la representación ilustra la interacción mediada por los genes Lr34, Lr46 y Lr67. Estos genes controlan una respuesta de la planta que retarda el desarrollo del patógeno, reduciendo la velocidad de aparición de las pústulas de la roya, lo que resulta en un menor número de lesiones. La resistencia mediada por estos genes no solo es efectiva contra P. triticina sino contra otras 2 especies de patógenos causantes de roya (Pucciniastriiformis f. sp. tritici y Puccinia graminis f. sp. tritici) y contra el mildiu del trigo (Blumeria graminis f. sp. tritici). Los genes de resistencia vertical y horizontal descritos en el caso de la roya solo se expresan durante la adultez de la planta. Para la resistencia horizontal, en los 2 ejemplos usados, la población del patógeno no está dividida en razas porque no existen interacciones específicas entre el patógeno y las variedades de la planta. Las variedades del hospedero, en cambio, exhiben diferentes respuestas al patógeno que resultan en diferentes niveles de enfermedad. En el caso de la resistencia vertical, solo existen 2 opciones: ausencia o presencia de la enfermedad.
Resistencia vertical: cuando las variedades del hospedero interactúan de manera diferencial o específica con las razas del patógeno.
Resistencia horizontal: cuando la interacción entre razas del patógeno y el hospedero no es específica.
En este contexto, las razas del patógeno existen porque interactúan de manera específica con variedades del hospedero de la planta en un evento de resistencia vertical, mientras que en la resistencia horizontal no existen razas fisiológicas. La resistencia horizontal está determinada por la presencia y acción de muchos genes, mientras la vertical es conferida básicamente por la presencia de un solo gen R. Esta característica hace que la resistencia sea superada por un cambio de raza del patógeno.
Teóricamente, la resistencia vertical es capaz de disminuir el inóculo efectivo para iniciar una epidemia (al no permitir la reproducción de una raza del patógeno en particular), pero no disminuye la tasa de reproducción de otras razas del patógeno. La resistencia horizontal, por el contrario, usa un mecanismo general: no es específica contra una raza en particular, sino que disminuye la tasa de reproducción para todas las razas56,57.
La capacidad del patógeno para iniciar una enfermedad se denomina patogenicidad y Van der Plank clasificó esta patogenicidad en 2clases: virulencia y agresividad. El término «virulencia» describe las razas del patógeno específicas para un cultivar, mientras que «agresividad» describe la capacidad del patógeno para infectar cualquier cultivar. Es válido recalcar que, según Van der Plank, solo se puede hablar de virulencia cuando hay razas del patógeno, esto es, cuando algunos aislados son capaces de infectar unos cultivares y otros no. En cambio, cuando una cepa del patógeno ataca a todos los cultivares hablamos de agresividad.
Según Van der Plank, virulencia y agresividad están negativamente correlacionadas, lo que significa que la especialización de una raza patogénica para infectar con gran éxito un determinado cultivar disminuye la capacidad para infectar otros cultivares y sobrevivir. Por otra parte, un patógeno capaz de infectar todos los cultivares tiene mayores chances de sobrevivir, pero, en general, menores probabilidades de infectar con éxito cada cultivar individualmente.
El concepto de gen por gen de Flor está estrechamente relacionado con el concepto de Van der Plank de resistencia vertical. El concepto de gen por gen describe la mayoría de las interacciones mediadas por reacción hipersensible (presencia de genes R y Avr) y es usado en muchos casos como sinónimo de resistencia vertical. Toda aquella resistencia no mediada por la reacción hipersensible es considerada resistencia horizontal. Esta definición de resistencia horizontal es limitada y solo describe un fenotipo de reacción de una planta individual que permite frenar de forma parcial el crecimiento del patógeno (resistencia parcial). Sin embargo, existen otras definiciones de resistencia horizontal que reflejan mejor sus características relevantes y constituyen una aproximación más útil para fines didácticos y de las actividades de mejoramiento.
En términos prácticos, la resistencia horizontal puede ser igualada a la denominada resistencia de campo (field resistance). Los mejores ensayos para la observación de la resistencia horizontal son los experimentos de campo11. La resistencia de campo es observada durante la evaluación de progenies o colecciones de cultivares en pruebas de campo, cuando se la considera una característica cuantitativa que exhibe una distribución continua. Se asume que esta resistencia es de carácter poligénico, porque la distribución continua de fenotipos de resistencia solo puede ser explicada por la combinación de varios loci genéticos (resistencia cuantitativa)8,41.
Aunque los conceptos de resistencia vertical y horizontal aún siguen siendo usados y generan confusión y controversia, los conceptos de reacción hipersensible y resistencia de campo son claros y aceptados sin discrepancias, porque reflejan las variaciones fenotípicas que es común observar en los ensayos de laboratorio y de campo. Además, estos términos son inherentemente agnósticos, sin ninguna presunción de mecanismos moleculares o requerimientos de hipótesis (conceptos o teorías) para ser definidos, o expectativas de posibles resultados.
Transformación del concepto de interacciones planta-patógenoLa fitopatología, como otras ciencias aplicadas de la biología, fue en un principio una disciplina en sí misma y las teorías antes expuestas definieron durante décadas su marco conceptual. Sin embargo, y al igual que otros campos del conocimiento, la fitopatología ha estado sujeta a la intensa interdisciplinariedad que ha caracterizado a esta última década, con la incursión de investigadores y conceptos provenientes de otras áreas, en especial de la ecología, la evolución y la biología molecular. Esta nueva generación de científicos no reconoce los límites previos de la disciplina y por eso es frecuente que surjan dificultades en la definición de conceptos y, a partir de ello, discrepancias respecto de cómo abordar problemas de investigación52. Podríamos decir que existe una convivencia de la fitopatología clásica con el enfoque interdisciplinario de las interacciones planta-patógeno y, en su forma más amplia, de las interacciones planta-microorganismo. Este último incluye la simbiosis entre plantas y micorrizas arbusculares, la fijación de nitrógeno por los nódulos radiculares resultantes de la interacción entre leguminosas y rizobios, y las relaciones planta-patógeno15.
Múltiples sinónimos han sido usados en las últimas 5décadas para explicar lo que Van der Plank llamó resistencia vertical y horizontal. La resistencia vertical de Van der Plank ha sido denominada resistencia monogénica (o conferida por unos pocos genes), resistencia de genes mayores, resistencia cualitativa, resistencia asociada a genes R, resistencia racial y resistencia mediada por reacción de hipersensibilidad. La resistencia horizontal, a su vez, ha sido llamada resistencia poligénica, resistencia asociada a genes menores, resistencia cuantitativa o asociada a quantitative trait locus (QTL), resistencia de campo o tolerancia14,33,34,38,42.
Cada denominación tenía como propósito diferenciar con claridad en cada caso específico el tipo de resistencia que ofrece una planta con la nueva información disponible. La confusión asociada a la división de la resistencia a patógenos en vertical y horizontal fue manifestada incluso en los inicios, cuando esta división fue propuesta34. Sin embargo, la idea tuvo más seguidores que detractores y aún hoy es usada con frecuencia. En la siguiente sección revisaremos los términos equivalentes a los de la clasificación clásica de resistencia vertical/resistencia horizontal desde el enfoque actual, con el propósito de crear un lenguaje común que ayude a comprender los avances más recientes en materia de resistencia de las plantas a los patógenos.
Resistencia cuantitativa y cualitativaLas respuestas de resistencia a las enfermedades en las plantas son complejas y contienen múltiples capas de interacción entre genes, proteínas y metabolitos. Como ya se indicó, tradicionalmente la resistencia ha sido clasificada en resistencia vertical (mediada por genes R) y horizontal (resistencia incompleta). Sin embargo, la nueva evidencia aportada indica que la resistencia horizontal y vertical no son más que un continuo de diferentes mecanismos desarrollados por la planta para interactuar con el ambiente externo, incluyendo los patógenos.
A continuación, presentaremos los mecanismos básicos de la interacción de las plantas con los patógenos. Algunos autores denominan a estos el nuevo «dogma central de la fitopatología»4:
Reconocimiento de los patrones moleculares asociados a los microbios (MAMP). Los MAMP son elicitores microbianos genéricos (podríamos llamarlos señales o huellas dactilares de los microorganismos), como la quitina y la flagelina, que pueden ser reconocidos por los receptores de reconocimiento de patrones (PRR) ubicados en la superficie celular de la planta. Si los PRR reconocen la presencia de MAMP en la superficie celular, se inicia una cascada de señales que conduce a una respuesta de resistencia débil y al desarrollo de inmunidad activada por microorganismos. El mecanismo de reconocimiento de los MAMP por los PRR no es único de las plantas y está presente en los animales, desde los insectos hasta los humanos46. Los PRR suelen contener o están asociados a cinasas sin arginina ni aspartato (non-RD)31,47.
Algunos patógenos son capaces de superar este nivel de resistencia de la planta secretando proteínas efectoras que interfieren con el metabolismo del hospedero y promueven su virulencia. Pero, a la vez, las plantas han desarrollado la inmunidad activada por efectores (ETI) para reconocer estos efectores o sus acciones. Esta respuesta conduce a la resistencia completa, sin producir ningún síntoma de la enfermedad. Las proteínas en la planta capaces de reconocer los efectores son las llamadas proteínas R (las asociadas a los genes R de Flor y Van der Plank). Estas proteínas R son, en su mayoría, del tipo receptores abundantes en leucinas con sitios de unión a nucleótidos (NBS-LRR)18. Los NBS-LRR y muchas de las proteínas R son capaces de unirse a productos de los genes Avr (los mismos genes de avirulencia propuestos por Flor)46.
A continuación, presentaremos una descripción de 2modelos de resistencia en plantas para explicar la información disponible: el modelo de zigzag de inmunidad vegetal y el modelo de invasión. Ambos están basados en los conceptos de inmunidad activada por patógenos (PTI) y ETI24,54.
Modelo zigzag de inmunidad vegetalEl modelo zigzag de inmunidad vegetal propuesto por Jones y Dangl23 identifica 2niveles en el sistema inmunitario de las plantas. El primer nivel es denominado inmunidad activada por patrones (PTI) y es capaz de reconocer las moléculas altamente conservadas que poseen la mayoría de los microorganismos (antes definidas como PAMP o MAMP). El segundo nivel es denominado ETI y es capaz de reconocer y responder a los factores de virulencia denominados efectores. Los efectores, de no ser reconocidos por la planta, son capaces de suprimir la respuesta del primer nivel y así facilitar el proceso de infección y colonización. Este es un modelo conceptual sencillo que explica la intrínseca red de procesos de la inmunidad vegetal, pero no tiene ninguna aplicación práctica porque no representa los aspectos mecanísticos de la interacción7,23,43.
Modelo de invasiónEn el modelo de invasión, los MAMP y los efectores son parte de los patrones de invasión (IP) que pueden ser reconocidos por la planta (en general, los IP son moléculas producidas directamente por el patógeno). La planta no solo reconoce los patrones de invasión, sino que también reconoce las moléculas resultantes del daño producido por la infección del patógeno, es decir, los patrones moleculares asociados al daño (DAMP). En resumen, en el modelo de invasión, las plantas reconocen 2tipos de patrones: los producidos por el patógeno (IP) y los resultantes de la infección por el patógeno (DAMP). Los IP son reconocidos por los receptores activados por IP (IPTR). Los genes R son IPTR activados por los efectores, que actúan como IP18.
En el contexto del modelo zigzag y del modelo de invasión, la resistencia cuantitativa (horizontal) y la cualitativa (vertical) pueden ser explicadas como un continuo dentro de un solo modelo conceptual18. La variación observada en la resistencia horizontal es explicada por variaciones cuantitativas en los componentes asociados a PTI y ETI, así como a otros mecanismos de rutas redundantes no asociados estrictamente a la resistencia a patógenos18,40. La figura 3 reúne la descripción de ambos modelos en un solo modelo consenso, en el que las primeras respuestas de la planta están asociadas a PAMP y, después, a efectores capaces de suprimir la respuesta de la planta. Sin embargo, esto no significa que algunos efectores no estén implicados en el reconocimiento de PAMP20,25,26,54,61.
 e inmunidad activada por efectores (ETI). La PTI está asociada a los patrones moleculares asociados a patógenos (PAMP). Ejemplos de PAMP son la flagelina de las bacterias y la quitina de los hongos. Los PAMP son moléculas estructurales comunes a varios géneros de patógenos, mientras los efectores son específicos para cada especie. Los efectores son proteínas implicadas en el desarrollo de una enfermedad con capacidad para suprimir la inmunidad de las plantas, o de producir susceptibilidad activada por (ETS). A) Resistencia completa al patógeno evitando la entrada a las células, o controlando la colonización de la célula por mecanismo de reacción hipersensible. B) Curva de las respuestas de defensa de la planta a través del tiempo. El patógeno inicia el proceso de infección y busca penetrar en las células vegetales. Durante este proceso la quitina (como ejemplo de PAMP) producida por el patógeno es reconocida por receptores superficiales de la célula (RCP1), que inician una respuesta de defensa por la planta que detiene el avance del patógeno. Sin embargo, el patógeno es capaz de cubrir sus rastros usando un efector (E) que evita el reconocimiento de la quitina y permite al patógeno avanzar en el proceso de infección. Nuevamente, la planta ha desarrollado mecanismos para reconocer este efector. RCP2 (otro receptor superficial de la célula) reconoce al efector e inicia un nuevo proceso de defensa. Estos procesos de defensa asociados directamente a un PAMP son denominados PTI. En el gráfico, los términos E1, E2 y E3 describen efectores. La susceptibilidad asociada a estos efectores (ETS) es la resultante de la supresión de la respuesta inmune de la planta por el patógeno. La resistencia completa (vertical) al patógeno puede ser alcanzada con cualquiera de los mecanismos de defensa, bien sea en respuesta a PAMP (PT1-PT2) o en respuesta a efectores (ET1-ET2). En múltiples ocasiones, la respuesta de defensa de la planta no puede controlar por completo al patógeno y resulta en resistencia parcial (horizontal). La curva roja describe la respuesta esperada en un proceso de resistencia vertical y la azul, la esperada durante la resistencia horizontal. C) Infección exitosa en un cultivar susceptible. Los mecanismos de defensa de la planta han fallado y ha dado como resulta la colonización exitosa de las células24,54.")
Descripción de la inmunidad activada por patrones asociados a patógenos (PTI) e inmunidad activada por efectores (ETI). La PTI está asociada a los patrones moleculares asociados a patógenos (PAMP). Ejemplos de PAMP son la flagelina de las bacterias y la quitina de los hongos. Los PAMP son moléculas estructurales comunes a varios géneros de patógenos, mientras los efectores son específicos para cada especie. Los efectores son proteínas implicadas en el desarrollo de una enfermedad con capacidad para suprimir la inmunidad de las plantas, o de producir susceptibilidad activada por (ETS). A) Resistencia completa al patógeno evitando la entrada a las células, o controlando la colonización de la célula por mecanismo de reacción hipersensible. B) Curva de las respuestas de defensa de la planta a través del tiempo. El patógeno inicia el proceso de infección y busca penetrar en las células vegetales. Durante este proceso la quitina (como ejemplo de PAMP) producida por el patógeno es reconocida por receptores superficiales de la célula (RCP1), que inician una respuesta de defensa por la planta que detiene el avance del patógeno. Sin embargo, el patógeno es capaz de cubrir sus rastros usando un efector (E) que evita el reconocimiento de la quitina y permite al patógeno avanzar en el proceso de infección. Nuevamente, la planta ha desarrollado mecanismos para reconocer este efector. RCP2 (otro receptor superficial de la célula) reconoce al efector e inicia un nuevo proceso de defensa. Estos procesos de defensa asociados directamente a un PAMP son denominados PTI. En el gráfico, los términos E1, E2 y E3 describen efectores. La susceptibilidad asociada a estos efectores (ETS) es la resultante de la supresión de la respuesta inmune de la planta por el patógeno. La resistencia completa (vertical) al patógeno puede ser alcanzada con cualquiera de los mecanismos de defensa, bien sea en respuesta a PAMP (PT1-PT2) o en respuesta a efectores (ET1-ET2). En múltiples ocasiones, la respuesta de defensa de la planta no puede controlar por completo al patógeno y resulta en resistencia parcial (horizontal). La curva roja describe la respuesta esperada en un proceso de resistencia vertical y la azul, la esperada durante la resistencia horizontal. C) Infección exitosa en un cultivar susceptible. Los mecanismos de defensa de la planta han fallado y ha dado como resulta la colonización exitosa de las células24,54.
En lo que respecta al patógeno, los efectores (que son reconocidos por las proteínas R) son polimórficos en la población; esto es, existen diferencias en un mismo efector para los individuos dentro de la población del patógeno. Estas diferencias son las responsables de las variaciones en la población del patógeno que conocemos como razas. Los receptores de las plantas también son polimórficos y esta variación es la que se observa como diferenciales en el caso de la planta. Entonces, tenemos una variación polimórfica para los genes de resistencia (R) y para los genes de avirulencia (Avr) específica para una interacción entre el patógeno y una especie de planta en particular, pero también mecanismos comunes de defensa a todas las plantas (y animales) mediados por MAMP (por ejemplo, la percepción de fragmentos de quitina provenientes de los hongos)18,46.
¿Cómo se mide la resistencia cuantitativa?La resistencia cuantitativa es mediada por varios genes que suelen encontrarse asociados con una región genómica o QTL. Cada uno de estos genes contribuye al efecto final sobre el fenotipo de resistencia al patógeno, es decir, tienen un efecto aditivo. Este efecto puede ser extrapolado a la combinación de varios QTL y generar resistencia completa, imitando el efecto de la resistencia vertical. Algunos QTL exhiben resistencia específica para un aislado. Estos casos son contrarios a la definición de eventos discretos que Van der Plank otorga a la resistencia horizontal y vertical. Sin embargo, tales definiciones son posibles si la resistencia horizontal y vertical son consideradas un continuo. A nivel molecular, los mecanismos asociados a la resistencia cuantitativa están codificados por genes con diferentes funciones: morfología, metabolismo o desarrollo de la planta, defensa basal, reforzamiento de la pared celular, detoxificación, transducción de señales de defensa y genes R alterados o débiles (NB-LRR) (estos últimos son parte de la PTI y la ETI)27,42.
Resistencia de campoLa resistencia de campo se mide aún de la misma manera que se medía hace 50 años, cuando se denominaba resistencia horizontal. La expresión fenotípica de la resistencia cuantitativa en ensayos de campo se denomina resistencia de campo y es la que le interesa al mejorador, al fitopatólogo y a los agricultores. La expresión genética es la que le interesa al genetista y, en combinación con las características del patógeno, la que es de interés para la biología evolucionaria. En el campo, la resistencia de un hospedero (progenies, variedades, líneas avanzadas o cultivares) es estimada a través de varios ciclos de la enfermedad usando los siguientes parámetros37:
Incidencia de la enfermedad: proporción de plantas (o unidades muestrales) infectadas, comúnmente medida como porcentaje de plantas enfermas.
Severidad (o intensidad) de la enfermedad: proporción de tejido afectado respecto del área total.
Cuando estas medidas son efectuadas a través del tiempo, son integradas en un solo valor estimado como el área bajo la curva (o la escalera) de la enfermedad27,50.
La resistencia también puede ser medida durante un solo ciclo de desarrollo de la enfermedad (infección monocíclica) en plantas inoculadas en condiciones de invernadero o campo. En estas condiciones, la medición de la resistencia está orientada a cuantificar la dinámica de los procesos infecciosos. Algunos de los parámetros usados son los siguientes: frecuencia de infección (proporción de esporas que producen lesiones esporulantes), período latente (tiempo transcurrido entre la infección y la producción de esporas), producción de esporas (esporas producidas por lesión o superficie de tejido en un intervalo de tiempo dado) y período de esporulación3,19,21,28,30,39,49,51,58,59.
La resistencia de campo y las mediciones en invernadero son estimaciones de la denominada resistencia cuantitativa de un hospedero frente al patógeno. En muchas ocasiones, durante el desarrollo de las evaluaciones de resistencia, es aconsejable introducir cultivares resistentes o susceptibles de fácil consecución y comunes a otros experimentos, que permitan comparar los resultados obtenidos con los de experimentos previos59. En algunos casos, la resistencia de los cultivares está basada en escalas desarrolladas para estimar su resistencia, independientemente del ambiente en el cual se efectuó la prueba. Estas escalas pueden estar basadas en evaluaciones visuales de cualquiera de los 3maneras de medir ya descritas3,19,21,28,30,39,49,51,58,59.
ConclusionesLa resistencia de las plantas a los patógenos fue clasificada en vertical y horizontal, y múltiples sinónimos de estos términos han sido creados con el correr del tiempo. En la última década, esos términos fueron reemplazados por los de resistencia cuantitativa y cualitativa. La resistencia cualitativa está mediada por los genes de resistencia R y su interacción con genes específicos del patógeno: los genes de virulencia. Este es el antiguo concepto de interacción gen por gen de Flor.
La evidencia molecular indica que no hay diferencias entre la resistencia cuantitativa y la cualitativa, sino que se trata de un continuo de respuestas de la planta al ambiente incluyendo los patógenos. Los patógenos activan principalmente 2sistemas de monitoreo de la planta: 1) el reconocimiento de los patrones moleculares asociados a los microbios (MAMP) y 2) la inmunidad activada por efectores (ETI), mecanismo capaz de reconocer las proteínas efectoras del patógeno o las modificaciones producidas en la célula hospedera por estos. Esta última es la que se conoce como resistencia vertical, porque genera fenotipos de respuesta bien definidos que pueden identificarse con facilidad.
La resistencia cuantitativa describe una distribución continua de la resistencia del hospedero a una enfermedad. Esta resistencia es frecuente observarla en pruebas de campo como la variación en incidencia o severidad de la enfermedad en una población de la planta. La resistencia cuantitativa evaluada en pruebas de campo se conoce como resistencia de campo.
FinanciaciónOBF compiló información y escribió este manuscrito como investigador de CORPOICA durante el desarrollo del proyecto C05727 (Tres accesiones de algodón del banco de germoplasma con resistencia a formas altamente virulentas de antracnosis (Colletotrichum gossypii-cephalosporioides) validadas en campo) financiado por el Instituto Colombiano Agropecuario (ICA) - Banco de Germoplasma. CORPOICA a partir de junio del 2018 cambió su identificación legal a AGROSAVIA.
Conflicto de interesesEl autor declara no tener ningún conflicto de intereses.
