




Although Cr(VI)-reducing and/or tolerant microorganisms have been investigated, there is no detailed information on the composition of the microbial community of the biocathode microbial fuel cell for Cr(VI) reduction. In this investigation, the bacterial diversity of a biocathode was analyzed using 454 pyrosequencing of the 16S rRNA gene. It was found that most bacteria belonged to phylum Proteobacteria (78.8%), Firmicutes (7.9%), Actinobacteria (6.6%) and Bacteroidetes (5.5%), commonly present in environments contaminated with Cr(VI). The dominance of the genus Pseudomonas (34.87%), followed by the genera Stenotrophomonas (5.8%), Shinella (4%), Papillibacter (3.96%), Brevundimonas (3.91%), Pseudochrobactrum (3.54%), Ochrobactrum (3.49%), Hydrogenophaga (2.88%), Rhodococcus (2.88%), Fluviicola (2.35%), and Alcaligenes (2.3%), was found. It is emphasized that some genera have not previously been associated with Cr(VI) reduction. This biocathode from waters contaminated with tannery effluents was able to remove Cr(VI) (97.83%) in the cathodic chamber. Additionally, through use of anaerobic sludge in the anodic chamber, the removal of 76.6% of organic matter (glucose) from synthetic waste water was achieved. In this study, an efficient biocathode for the reduction of Cr(VI) with future use in bioremediation, was characterized.
Aunque se ha investigado sobre los microorganismos reductores y/o tolerantes de Cr(VI), no hay información detallada sobre la composición de la comunidad microbiana del cátodo de una Celda de Combustible Microbiana para la reducción de Cr(VI). En esta investigación se analizó la diversidad bacteriana de un biocátodo usando pirosecuenciación 454 del gen 16S rRNA. Se encontró que la mayoría de las bacterias pertenecieron a los filos Proteobacteria (78,8%), Firmicutes (7,9%), Actinobacteria (6,6%) y Bacteroidetes (5,5%), comúnmente presentes en ambientes contaminados con Cr(VI). Se encontró como género dominante a Pseudomonas (34,87%), seguido por los géneros Stenotrophomonas (5,8%), Shinella (4%), Papillibacter (3,96%), Brevundimonas (3,91%), Pseudochrobactrum (3,54%), Ochrobactrum (3,49%), Hydrogenophaga (2,88%), Rhodococcus (2,88%), Fluviicola (2,35%) y Alcaligenes (2,3%). Se destaca que algunos géneros no han sido previamente asociados con la reducción de Cr(VI). Este biocátodo procedente de aguas contaminadas con efluentes de curtiembres fue capaz de remover Cr(VI) (97,83%) en la cámara catódica. Adicionalmente, a través del uso de lodo anaeróbico en la cámara anódica, se logró la remoción del 76,6% de materia orgánica (glucosa) a partir de agua residual sintética. En este estudio se caracterizó un eficiente biocátodo para la reducción de Cr(VI) con futuro uso en biorremediación.
In a biocatalytic microbial fuel cell (MFC) anodic reactions (the transfer of electrons from the microorganism to the anode) are used for the production of current while the cathodic reactions (transfer of electrons from the cathode to the microorganism) have been recently applied for current consumption8. The biocathodes may be a better alternative to abiotic catalysis, because the bacteria are used as biocatalysts on low cost carbon cathodes16 and are able to accept electrons and reduce compounds which, in their oxidized form, are potential environmental pollutants, such as Cr(VI), NO3−, chloroethenes, 2-chlorophenol, ClO4−, U(VI) and CO217.
Recently, biocatalytic MFCs have shown promising results for the reduction of Cr(VI) to less toxic Cr(III)43. These MFCs work by electron transfer from cathodes to electrochemically active bacteria, and have received more attention for the removal of Cr(VI) due to their self-sustainability, low cost, low maintenance and minimal sludge generation48. The results of the investigations demonstrate the removal of Cr(VI) using microorganisms from domestic sewage42, anaerobic digesters38, polluted soil18, primary clarifier effluent from wastewater treatment plants17, anaerobic digester sludge46, and the cultivation of the bacteria Shewanella oneidensis MR-18,47. These investigations demonstrate the potential of microorganisms to reduce Cr(VI), it is therefore important to explore different sources of microorganisms in order to obtain more efficient biocatalysts for Cr(VI) reduction. In previous studies34 the waters of the Pasto River, which present contamination with effluents from artisan tanneries, were used as inoculum, achieving the removal of Cr(VI), however the composition of the microbial community that might contribute to this purpose is unknown.
In addition, little research aimed at studying the diversity of this type of microbial communities has been conducted, which does not allow for wide discussion about its structure and composition. Tandukar et al.38 studied the bacteria that colonize the cathode of a Cr(VI)-conducting MFC and obtained RFLP profiles of the 16S rRNA gene from randomly selected clones, their sequencing revealed the dominance of phylogenetically closely related to Trichococcus pasteurii and Pseudomonas aureginosa, putative reducers of Cr(VI). Another investigation was realized by Li et al.25, who performed a RAPD analysis for cathode biofilms of single compartment MFCs with Cr(VI), Fe(III) and O2 as terminal electron acceptors (TEAs), demonstrating that cathode bacteria vary according to the different TEAs and that in order to facilitate the identification of bacteria, a microbial community analysis based on the 16S rRNA gene must be performed. Wu et al.46 analyzed the microbial community of a Cr(VI) reducing biocatalyst using PCR-DGGE, showing through DGGE profiles that γ-Proteobacteria was the most abundant division. Recently Xue et al.48 isolated four electrochemically active bacteria from Cr(VI)-reduced biocathode MFCs, classified as Stenotrophomonas sp. YS1, Stenotrophomonas maltophilia YS2, Serratia marcescens YS3, and Achromobacter xylosoxidans YS8. The investigation was conducted employing, for the first time, a fluorescent Cr(III) probe, with a naphthalimide-rhodamine base, in order to perceive Cr(III) ions. The investigation showed that although Cr(VI) is reduced to Cr(III), forming Cr(OH)3 and Cr2O3 precipitates on the surface of cathodic electroactive bacteria, there is, at this point in time, no information about the occurrence of intracellular Cr(VI) reduction.
The diversity of these microbial communities will be influenced by different factors such as the type of inoculum and environment from which it was extracted, pH, temperature, types of substrates, and cell design. It has been shown that different electrode materials, surface area, and porosity, have an impact on microbial adhesion and growth, although the effect in the cathode on microbial composition is still unknown36. As mentioned above, it is inferred that the microbiota can become widely diverse in its structure and composition.
The objective of the present investigation was to analyze the bacterial diversity of the biofilm associated with the cathode of a Cr(VI) reducing MFC which uses a low cost separator that is salt bridge (SB). The pyrosequencing 454 of the V1–V3 hypervariable regions of the 16S rRNA was used for the analysis. This research offers new knowledge about the bacterial diversity of the biofilm formed by bacteria from the Pasto River, contaminated with affluents from artisan tanneries. In addition, the elimination of organic matter in the anodic chamber of the cell was evaluated.
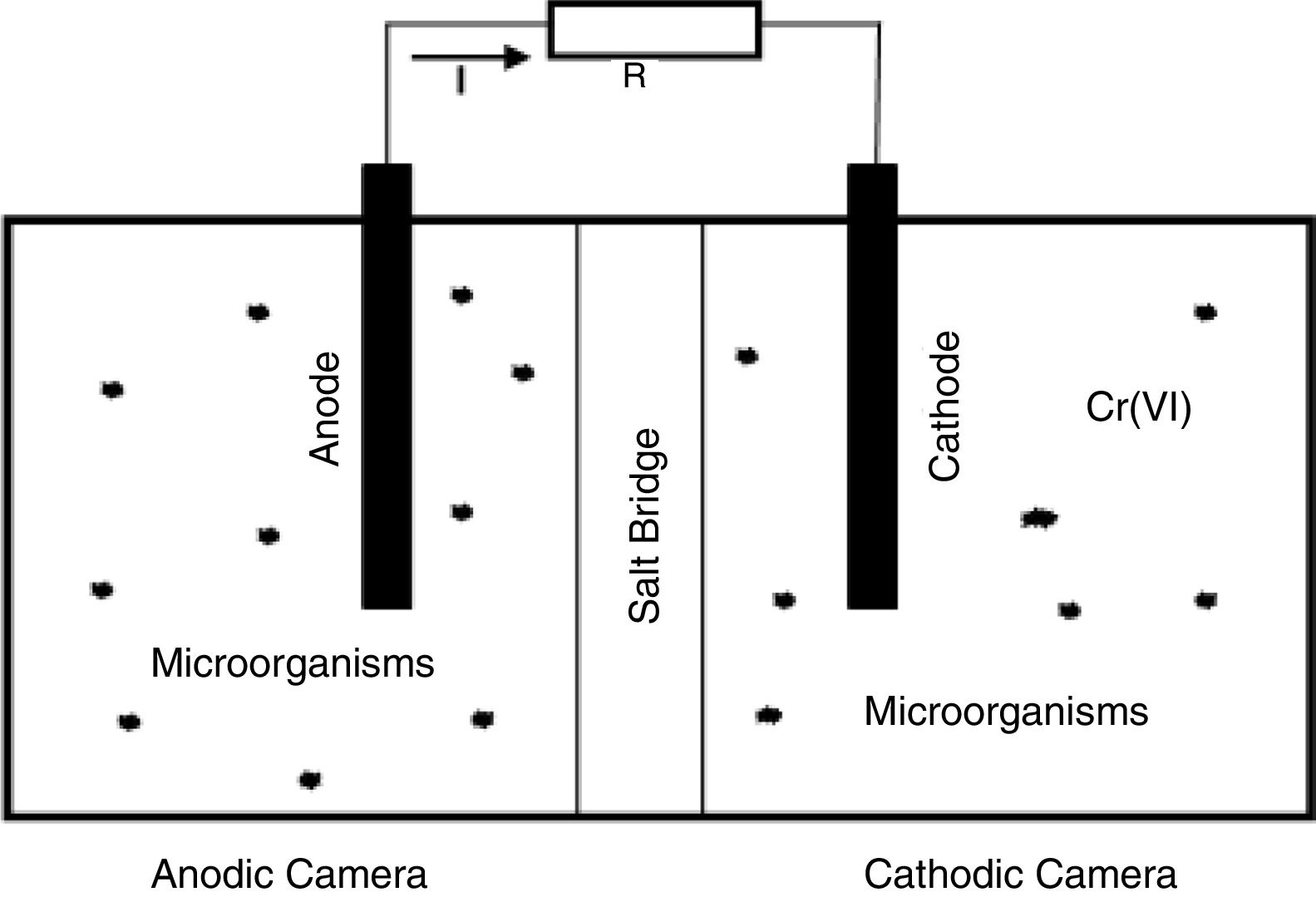
Materials and methodsMFC configurationThis study was conducted using a MFC-SB, built with low-cost materials, and consisting of a double chamber system (two rectangular acrylic compartments with volumes of 0.3l), separated by a salt bridge (1.5% agar, Oxoid, and 0.25M KCl electrolyte solution, Merck). Cylindrical graphite bars were used as electrodes (7cm long×0.7cm in diameter), along with a rectangular graphite bed (6.6cm×6cm) for a total contact area of 45cm2. Both the anode and cathode were connected by attaching a conducting wire to an external resistor of 500Ω (Fig. 1).
Inoculation and operating conditions.")
The anode chamber of the MFC-SB was inoculated with 10% of anaerobic sludge from the solid waste treatment plant of the city of Pasto (Antanas Sanitary Landfill), while the cathodic chamber of the MFC-SB was inoculated with 10% of Pasto River water contaminated with wastewater from tanneries in the city of Pasto. The inoculated samples were collected and maintained at room temperature until the time of use.
For the anodic chamber, 300ml of synthetic wastewater was used in the manner described by Cha et al.5, modified at the carbon source using, in this case, 0.1M acetate (Sigma Aldrich) (g/l): KH2PO4 (8.2), K2HPO4 (7.0), NH4Cl (0.1), NaHCO3 (0.2), MgSO4·7H2O (0.01) and micronutrient solution (mg/l): CaCl2·6H2O (130), FeCl3 (1000), H3BO3 (6), ZnCl2 (70), CuCl2 (2), MnSO4·H2O (100), (NH4)6Mo7O24·4H2O (206), AlCl3 (50), CoCl2 (238), and NiCl2 (24). For the cathode chamber, 300ml of the macronutrient solution was employed, as described by Huang et al.18 (g/l): KH2PO4 (4.4), K2HPO4 (3.4), NH4Cl (1.3), NaCl (0.5), CaCl2 (0.0146), NaHCO3 (1.0), and micronutrients proposed by Cha et al.5 (substituting MgCl2 for MgSO4·7H2O), but was modified using 1ml/l of the micronutrient solution proposed by Rabaey et al.33 Additionally, the cathodic solution was supplemented with K2Cr2O7 (Sigma Aldrich) (Cr(VI)) at 61.3ppm.
Previously, the formation of biofilms was initiated on anodes and cathodes. For this reason, the electrodes were maintained under anaerobic conditions, at room temperature, in flasks containing the respective inoculum and culture medium. The bioelectrodes were then transferred to their respective chambers and the MFC-SB was operated in batch mode until constant voltages close to zero were observed. In the negative control group, a cell of the same characteristics was used and operated under the same conditions, using instead, electrodes without biofilm (previously sterilized).
Amplification and 454 pyrosequencing of the 16S rRNA geneAfter the operation time of the MFC-SB, the biocathode was washed several times with phosphate buffered saline (8.0g/l NaCl, pH 7, Merck) and with a sterile scalpel the entire biofilm was carefully removed, avoiding the addition of graphite2,38. The total DNA was extracted from a biofilm sample (approximately 0.25g), using the PowerSoil DNA Isolation kit (MO BIO, Laboratories, Inc.), increasing the time of centrifugation in the steps of cell lysis and separation of contaminants to 2min. The amount and quality of the DNA (∼25ng/μl) were assessed by electrophoresis using Lambda phage DNA (Fermentas) at concentrations of 5–50ng/μl and the recording was conducted using a Photo Documentor System (LabNet, Gentech). Pyrosequencing was performed using the total DNA obtained from the biofilm in the Molecular Research DNA (MR DNA, Texas, USA) laboratories, starting with the bTEFAP® process12, modified with universal primers (27F and 519R) to amplify the V1–V3 (∼492bp length) hypervariable regions of the 16S rRNA gene using the Hot Star Taq Plus Master Mix Kit (Qiagen, Valencia, CA). The following thermocycling conditions were employed: 94°C for 3min, followed by 28 cycles of 94°C for 30s; 53°C for 40s, 72°C for 1min; and a final elongation step at 72°C for 5min. All amplification products were mixed at the same concentrations and purified using Agencourt Ampure beads (Agencourt Bioscience Corporation, MA, USA). The samples were sequenced using Roche 454 FLX Titanium instrument and reagents, according to the manufacturer's instructions.
Processing of pyrosequencing data16S rRNA pyrosequencing data was processed using an analysis guide (www.mrdnalab.com, MR DNA, Shallowater, TX), removing barcodes and primers and then short sequences (<200bp). Sequences with ambiguous bases were also removed (Ns), and those with homopolymers greater than 6bp. After the denoising process, the Operational Taxonomic Units (OTUs) were defined by grouping the sequences at a divergence of 3% (97% similarity), removing singleton sequences and chimeras. The final OTUs were taxonomically classified using the BLAST algorithm, cross-referencing the databases Green Genes, RDPII and NCBI. Subsequently, the quality of the sequences was confirmed using the Quantitative Insights Into Microbial Ecology (QIIME) software10. Based on the abundance matrices of the bacterial species, the relative abundance percentage and alpha diversity indexes (Chao1, Shannon-Weinery Simpson) were calculated using Past software15.
Cr(VI) reduction in the MFC-SB cathode chamberTo evaluate the hexavalent chromium (%Cr(VI)) reduction, the APHA Method 3500-CR (Standard Methods for the Examination of Water and Wastewater), which is characterized by the formation of a colored complex with 1.5-diphenylcarbazide (Panreac) was used. Absorbance measurements were performed at 543nm on a spectrophotometer (Pharo 3000, Merck-Germany). Initially, a calibration line was constructed by preparing K2Cr2O7 solutions in a concentration range of 0.2–1ppm34. In the process the linear range, precision, accuracy, limit of quantification, and limit of detection were evaluated. The concentration of Cr(VI) remaining in the MFCs at the beginning and at the end of the period of operation was determined by interpolation.
Removal of organic matter in the anode chamber of the MFC-SBThe amount of organic matter removed in the MFC was evaluated by determining the Chemical Oxygen Demand (COD) at the beginning and end of the cell's operating period. The determination was performed using the APHA Method 5220 B COD (Standard Methods for the Examination of Water and Wastewater) and a digestion reactor (E&Q Thermoreactor). Data was expressed as percentage of organic matter removal (%OM). The calibration line was constructed by preparing solutions of the primary potassium acid phthalate (Panreac) in a concentration range equivalent to 100–900mgO2/l. Each point on the line was measured in triplicate and the results were expressed as the data±standard deviation.
MFC-SB electrical performanceInitially, the cells were operated in open circuit (without external resistance) until the voltage stabilized. The circuit was then closed, and voltage measurements were performed every 10min using a Data Acquisition System (DAQ). The operating time was set when the voltage dropped to a value close to zero. The current (I) generated in the MFC was calculated based on Ohm's law (I=V/R), where V is the voltage in volts (V) and R is the external resistance (500Ω). Based on these findings, the Power Density (PD) and Coulomb Efficiency (CE) were determined.
The PD that measures the power generated per unit area of the electrode was determined by the following expression1,36:
where V is the voltage, I is the current, and A is the area of the electrode (cm2).
The CE, which is defined as the ratio of the effective charge transferred from the substrate to the anode at the maximum possible load (assuming that all the substrate removed produces electric current), was determined according to the following expression1:
where Cp is the amount of charge calculated by the integration of the current in time and Cti is the theoretical amount of charge calculated by following expression1:where F is the Faraday constant (96.485C/mol of electrons), b is the number of moles of electrons produced per mole of substrate, s is the substrate concentration, V is the volume of the liquid and M is the molecular weight of the substrate.Results and discussionRichness and bacterial diversity of MFC-SB cathode biofilm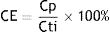
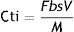
From the total MFC-SB cathode biofilm DNA, subjected to the 454 pyrosequencing process of the V1–V3 regions of the 16S rRNA gene, 8868 high quality reads (mean length 445bp) were obtained; that is 94.86% of the total reads prior to processing (9349). The high quality reads obtained were grouped into 124 OTUs at a distance of 3%. The total number of OTUs expected by the Chao1 estimator was 131, of which 124 (94.6%) were found, that is, pyrosequencing was able to capture most of the bacterial species present. When evaluating the alpha diversity of the MFC-SB, the indices of Shannon (3.48) and Simpson (0.90) were estimated. The latter evidences a high dominance, shown by the high abundance of one of its OTUs which exerts more control on the structure of the community, as will be later shown.
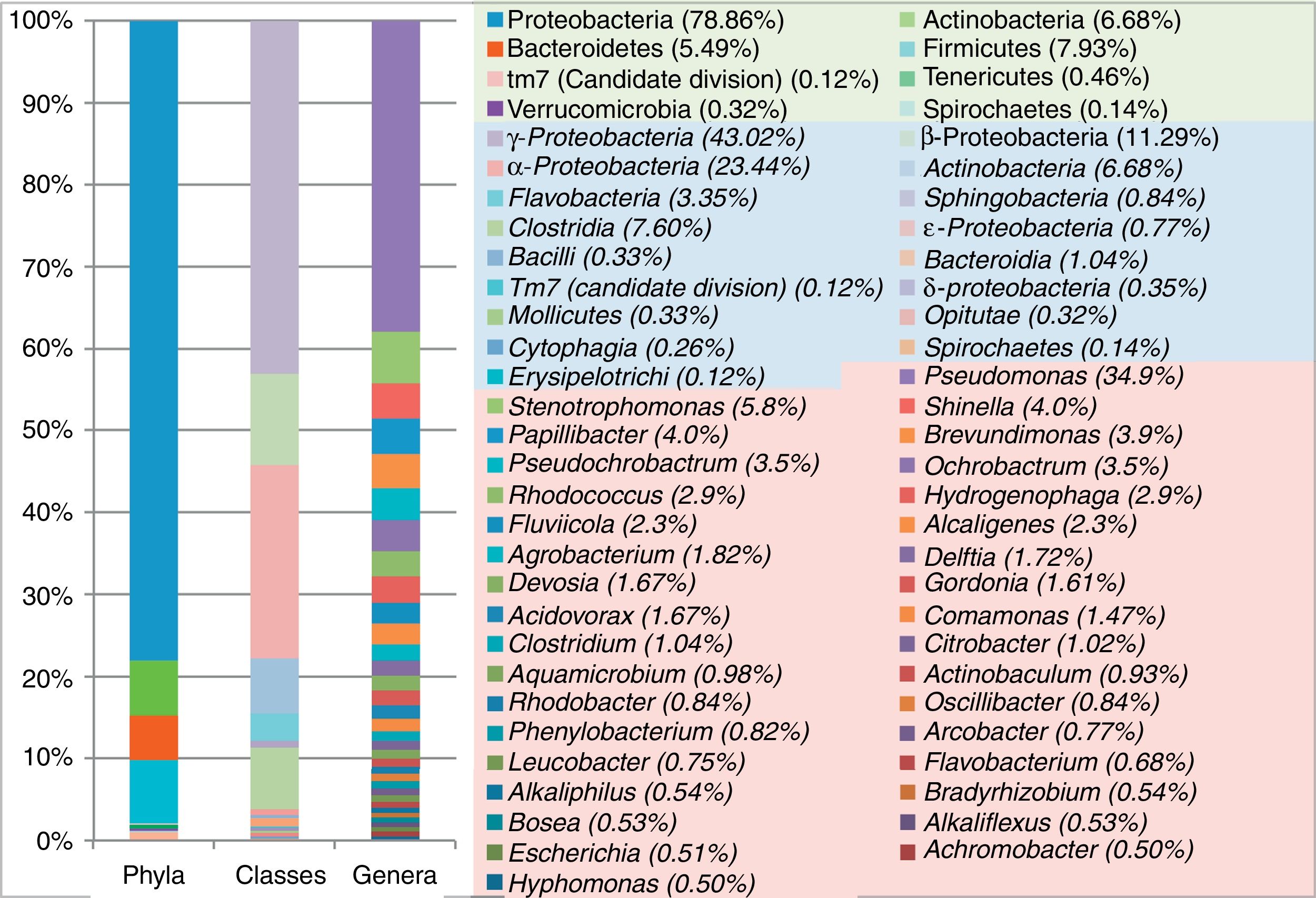
Analysis of the bacterial diversity of the MFC-SB biocathodeIn this investigation, the V1–V3 hypervariable regions of the 16S rRNA gene sequenced allowed a high taxonomic resolution. The diversity in the MFC-SB biocathode was represented by 7 phyla, the most abundant being Proteobacteria (78.8%), followed by phyla Firmicutes (7.9%), Actinobacteria (6.6%), Bacteroidetes (5%), and later four phyla with lower abundance of Tenericutes (0.4%), Verrucomicrobia (0.3%), Spirochaetes (0.1%) and candidate phylum TM7 (0.1%) (Fig. 2).

The most abundant phyla detected in MFC-SB (Proteobacteria, Actinobacteria, Firmicutes and Bacteroidetes) have also been previously reported as being more abundant in the bacterial community of an environment contaminated with Cr(VI)11. Phyla Verrucomicrobia and Spirochaetes occur in very low abundance, and no reports are known of their association with environments contaminated with Cr(VI). However, Sun et al.36 and Lesnik and Liu23 report them as representative at the cathode and anode of Cr(VI) non-reducing MFCs, different from those used in this study, though further studies are required in order to explain their presence in this MFC-SB. With regard to the 2 phyla of lower abundance, the presence of the chrA gene for Cr resistance has been reported in Tenericutes19, while TM7 has been detected in the biofilm of the anode produced from soil contaminated with polycyclic aromatic hydrocarbons (PAHs) from an aerated cathode cell50.
The abundance of the phylum Proteobacteria could be due to the diversity of its species being adapted to a wide variety of environments, including the one generated in MFC-SB, requiring the capacity to express a metabolism related to the reduction of Cr(VI). In this sense, a high percentage of OTUs belonged to the classes γ-Proteobacteria (43.02%), α-Proteobateria (23.44%) and β-Proteobacteria (11.29%) (Fig. 2). In a recent study based on DGGE analysis, Wu et al.46 found that the γ-Proteobacteria (bands 1, 4, 5, 7 and 13) were the most abundant, and demonstrated their function as electrotrophic bacteria on biocathodes to reduce Cr(VI) in MFCs; the researchers also discovered β-Proteobacteria (band 8) and α-Proteobateria (band 12) in the Cr(VI) reducing biocathode. It should also be mentioned that in the present investigation the remaining percentage of OTUs corresponded to the following classes: Clostridia, Actinobacteria, Flavobacteria, Sphingobacteria, Bacteroidial, ɛ-proteobacteria, δ-proteobacteria, Opitutae, Bacilli, Cytophagia, Mollicutes, Spirochaetes and Erysipelotrichi (Fig. 2).
The phylum Proteobacteria excelled by the detection of different bacterial genera (Fig. 2), as described below. The dominant genus Pseudomonas (34.9%) was detected in the MFC-SB, species of this genus have been used in different types of bioreactors to detoxify Cr(VI) and have been shown to be strong reducing agents of this contaminant by the high rates of reduction that it produces31. Its dominance in this type of environment can be attributed to its competence to develop various strategies to resist the harmful effects of Cr(VI), such as the expulsion of cytoplasm or its reduction to Cr(III) by the activity of the enzyme chromate reductase (ChrR), which stands out as one of the mechanisms of resistance to Cr(VI) and has been found in several species of the genus Pseudomonas: P. ambigua and P. putida31. In the MFC-SB biocathode, the following bacteria were detected: P. stutzeri, P. mendocina, P. pseudoalcaligenes, P. agarici, P. nitroreducens, P. synxantha, P. fragi, P. poae, P. putida and Pseudomonas sp. In previous studies, Tandukar et al.38 were able to characterize the species P. aeruginosa as a putative Cr(VI) reductant when using RFLP on the DNA obtained from the biocathode of a Cr(VI) reducing MFC. Wu et al.46 using DGGE analysis detected P. stutzeri (band 1) and P. extremorientalis (band 7) in the Cr(VI) reducing MFC biocathode. As a result, this investigation has enriched the information on the pseudomonas associated to the biocathodes of Cr(VI) reducing MFCs. The abundant Ochrobactrum (3.5%) genus (O. antrhopi, O. lupini and Ochrobactrum sp.) was detected, as well as O. antrhopi and O. tritici, which were isolated from environments contaminated with Cr(VI) and have been shown as potentially resistant and reducing agents of this metal, as they are carriers of chromate-resistance genes, located in transposons7,41. It has been shown that species such as Ochrobactrum sp. and P. aeruginosa are able to grow in Cr(VI)-contaminated environments and also produce exopolysaccharides (EPS) for biofilm formation21. This is likely to be an adaptive resistance strategy against metal, and both provide them with protection and acts as a form of communication.
With respect to the abundant genus Stenotrophomonas (5.8%) (S. maltophilia, S. acidaminiphila, S. rhizophila and S. nitritireducens), it has been determined that the species S. maltophilia and Stenotropohomonas sp., have high Cr(VI) reduction rates and the ability to produce EPS and biofilm formation on inert surfaces14,29,48. Morel et al.29 conclude that the higher EPS production and biofilms developed by Stenotrophomonas sp. exposed to low doses of Cr(VI) (100mg/l) can be increased to protect cells from Cr stress. In their investigation, Ge et al.14 showed that the strain Stenotrophomonas sp. D6 completely reduced Cr(VI) in a solution containing K2Cr2O7 at 200mg/l after 72h of incubation, and was tolerant to K2Cr2O7 at 1600mg/l, demonstrating that this strain was more effective than other strains of the genus in the reduction of Cr(VI). The authors also demonstrated that the presence of Cu2+, Fe3+ and Zn2+ increases Cr(VI) reduction while Ag+ and Co2+ inhibit their reduction in this strain. These results are relevant because they indicate that bacteria with similar characteristics to D6 may be present in the bacterial community characterized here and that they are affected by other metals present in the solution that must be determined in future studies.
The abundant Brevundimonas genus (3.9%) was represented by the species B. diminuuta, which has been found in high abundance in samples contaminated with Cr(VI)26. The following abundant genera were also detected in the MFC-SB biocathode: Pseudochrobactrum (3.5%) (P. saccharolyticum and Pseudochrobactrum sp.) and Alcaligenes (2.3%) (A. faecalis). It was recently identified for the first time that bacteria of the genus Pseudochrobactrum, reduces and tolerates Cr(VI), highlighting the species Pseudochrobactrum sp. B5 by tolerating concentrations of 2000mg/l and reducing 1000mg/l of Cr(VI) in 96h growing in LB medium13. Therefore, the presence of species of this genus in the MFC-SB biocathode is a relevant result and these bacteria could have equal or greater potential than those studied by the authors. Similarly, in previous studies it is recognized that the genus Alcaligenes participates in the intracellular reduction of Cr(VI), distinguishing A. eutrophus which uses the enzyme ChrA for the resistance and reduction of Cr through the ChrA enzyme39. In this study, its participation in the biocathode of a Cr(VI) reducing MFC is shown for the first time.
It is important to highlight the presence of abundant genera Shinella (4%) and Hydrogenophaga (2.9%) (H. palleronii). In the first genus, no previous reports have been found to associate it with Cr(VI). This may constitute the first report of bacteria of this genus that fulfill some function in the biocathode used for the reduction of Cr(VI). The genus Hydrogenophaga is known to have the ability to form biofilm on the anode of a MFC22 and was recently detected in a site contaminated by Cr(VI) and chlorinated ethenes30. In this investigation, their participation in the reduction of Cr(VI) in the MFC biocathode is discussed for the first time.
Other abundant genera detected in the MFC-SB biocathode were: Rhodococcus (2.9%) (R. globerulus, R. erythropolis and R. fascians) belonging to the phylum Actinobacteria (Fig. 2). Previous studies have shown that bacteria of this genus have the ability to reduce Cr(VI) – for example, Sun et al.35 demonstrated that the Rhodococcus Chr-9 strain has the ability to reduce Cr(VI) when glucose and LB were used as the carbon source. Patra et al.32 demonstrated that R. erythropolis reduces Cr(VI) in minimal medium, supplemented with glucose as the sole carbon source of carbon. This study demonstrates the presence of bacteria of this genus in the MFC-SB biocathode that could be involved in the reduction of Cr(VI) in a medium containing NaHCO3 as the carbon source. The abundant genus Papillibacter (4.0%) (Papillibacter spp.) has not been related to the reduction of Cr(VI) in previous studies, however, the phylum Firmicutes to which it belongs (Fig. 2), is highly abundant in soils contaminated with tannery effluents11. Similarly, the abundant genus Fluviicola (2.35%) (Fluviicola spp.) has not been associated with Cr(VI) reduction, although the phylum Bacteroidetes (Fig. 2) is present in environments contaminated with Cr(VI)46. In this study, we therefore discuss for the first time, bacterial species of these phyla with the potential to tolerate or reduce Cr(VI).
Of the 23 non-abundant genera (0.5–1.9%) detected in the MFC-SB biocathode (Fig. 2), 16 genera (Citrobacter, Clostridium, Delftia, Achromobacter, Agrobacterium, Escherichia, Leucobacter, Rhodobacter, Comamonas, Devosia, Gordonia, Alkaliphilus, Hyphomonas, Acidovorax, Bosea and Flavobacterium) have been reported in previous studies as potential Cr(VI) reducers3,4,6,19,28,37,39,40,44,45,49. While their role in Cr(VI) reduction is not yet known, the remaining 7 genera (Actinobaculum, Aquamicrobium, Oscillibacter, Phenylobacterium, Arcobacter, Bradyrhizobium and Alkaliflexus) their presence in the cathode biofilm of the MFC-SB could constitute a first report of their association with the reduction of this metal.
Evaluation of Cr(VI) removal in the MFC cathode chamberThe concentration of Cr(VI) at the beginning and end of the operation period in the MFC was determined according to the previously mentioned methodology. In the calibration process used to determine the Cr(VI), r2=0.9976 and a detection limit of 0.0088ppm were established. The initial concentration of Cr(VI) was 61.3ppm, resulting in a high% Cr(VI) of 97.83% (Table 1) in the cell process, demonstrating that these systems have a high Cr(VI) reduction capacity. In the analysis, a control cell with the same characteristics but without biofilm was used, and it was observed that after of the operation period, the device did not present any electrical response, demonstrating the activity of the microorganisms in the process of degradation of the acetate.
At the end of the operating period,%Cr(VI) (higher than 97%) was found in the cathode solutions of the MFC, showing that Cr(VI) is used by cathode microorganisms as an TEA, reducing it to Cr(III). This behavior has been recorded by different authors8,42, who postulate that the reduction possibly occurs through enzymatic mechanisms associated with the cell membrane or the cytoplasm, although the metabolic pathways of Cr(VI) reduction have not been fully elucidated. Most of the bacterial species detected in the MFC-SB cathode biofilm belong to genera which have been demonstrated as Cr(VI) resistant and/or -reducing in previous studies.
Evaluation of the removal of organic matter in the anode chamber of the MFCIn the same period of operation, an appreciable reduction of organic matter (76.6%) (Table 1) was also observed, which supports the idea that there is also microbial activity in the anode chamber9. Similar observations were made by Revelo et al.34, who evaluated the% OM at different operating times and found the highest percentages (91–97%) in cells operated over a longer period of time. In general, the organic matter removed in the anode is representative and comparable with data published in the scientific literature. For example, Cirik9 found high% OM (90%), demonstrating a high efficiency of the microbial activity in the anodic chamber, product of the adaptation of the microorganisms to the different conditions of the cell and to the assimilation capacity of the substrate used.
Evaluation of the electrical performance of the MFC-SBThe MFC-SB was operated in an open circuit for 7 days and closed circuit for 51 days. The evaluation of MFC-SB electrical performance in closed circuit shows that following indicators (Table 1): the highest voltage (8.033mV), PD (2.806E−06mW/cm2) and CE (0.044%). A high electrical potential was presented when the circuit was closed (27.041mV) because the anode is polarized by the effect of the accumulation of electrons released by oxidation of the organic matter. Once the external resistance between the anode and the cathode of the MFC (closed circuit) is connected, the accumulated electrons move, producing a low electric current. It was observed a voltage fluctuation that may be related to microbial activity in the anodic biofilm, where exoelectrogenic bacteria act, as well as to those that are not able to transfer electrons to the anode but degrade organic matter. According to Kiely et al.20, the combination of microbial populations is related to the products that are generated in the medium during the formation of the biofilm which affects on the generation of voltage, since the syntrophic processes are key in the exoelectrogenesis of a substrate.
The PD calculated was generally low compared to previous investigations38,47 in Cr(VI) reducing biocatalytic MFCs, which vary between 1.7mW/cm2 and 56mW/cm2. Similarly, the CE found in the cells was lower compared to other investigations in which CE was of 53%42. It should be noted that the antecedents that are cited use an proton exchange membrane (PEM) in this research using a salt bridge, which coincides with the results of Li et al.24, who proposes that PEM as a separator is more efficient than the salt bridge and offers better conditions for the mobility of electrons from the substrate to the anode. On the other hand, it is known that the electrical performance of a cell with salt bridge is low, however, we use this type of separator because of its low cost. Although a usable energy is not generated, a high removal of Cr(VI) was achieved.
In addition other factors such as MFC configuration, distance between electrodes, the presence of oxygen, nitrates and sulfates acting as TEAs27, and pathways such as methanogenesis, which consume more than 26% of total electrons38, can also explain the low electrical performance. These two last factors prevent the electrons produced in the anode chamber from being transferred to the anode, reducing the circulation of electrons by the external circuit, thus affecting the electrical performance38.
ConclusionsThe bacterial community characterized in the MFC-SB is composed by 4 abundant phyla (Proteobacteria, Actinobacteria, Firmicutes and Bacteroidetes). The greater control on the structure of the community was exerted by the genus Pseudomonas, though the abundant genera Ochrobactrum, Stenotrophomonas, Brevundimonas, Pseudochrobactrum, Alcaligenes, Hydrogenophaga, Rhodococcus, Shinella, Papillibacter and Fluviicola were highlighted. The last three genera have not been reported as reducing or tolerating Cr(VI), and future studies are required to understand its function in the biocathode reducer of this metal. Most of the bacterial species detected have the capacity for Cr(VI) reduction and/or tolerance, although the dominance of the genus Pseudomonas is emphasized. Cr(VI) reduction observed in the MFC-SB could be the result of the complex interaction among the bacterial species detected which should be further explored in future investigations. This research broadens our knowledge of the bacterial diversity associated with the Cr(VI) reducing MFC-SB biocathode, formed by bacteria from the waters of the Pasto River contaminated with effluents from artisan tanneries.
Under the mentioned conditions the energy generation was very low and it is therefore necessary to conduct an in-depth study of the factors that influence the electric performance of the cell. Nonetheless, it must be emphasized that in this investigation, using a low cost separator, an important removal efficiency was obtained, which can be attributed to the biofilm formed on the electrodes. The reduction to the less toxic form of chromium (trivalent chromium) and the reduction of the organic matter show a possible application of these systems in the treatment of wastewater, demonstrating the potential for bioremediation processes of contaminated effluents.
NoteThis document is the product of a multidisciplinary research, in which the bacterial diversity associated to the highly reducing biocathode of Cr (VI) of a Microbial Fuel Cell was analyzed, using a low cost separator (saline bridge), synthetic waste water and microorganisms involved in bioelectrochemical processes, coming from autochthonous sources. This research explores a technology potentially applicable to the bioremediation of wastewater.
Conflict of interestThe authors declare that they have no conflicts of interest.
The authors are grateful for the financial support provided by the Vice-Rectory of Postgraduate Research and International Relations - VIPRI, University of Nariño.
