





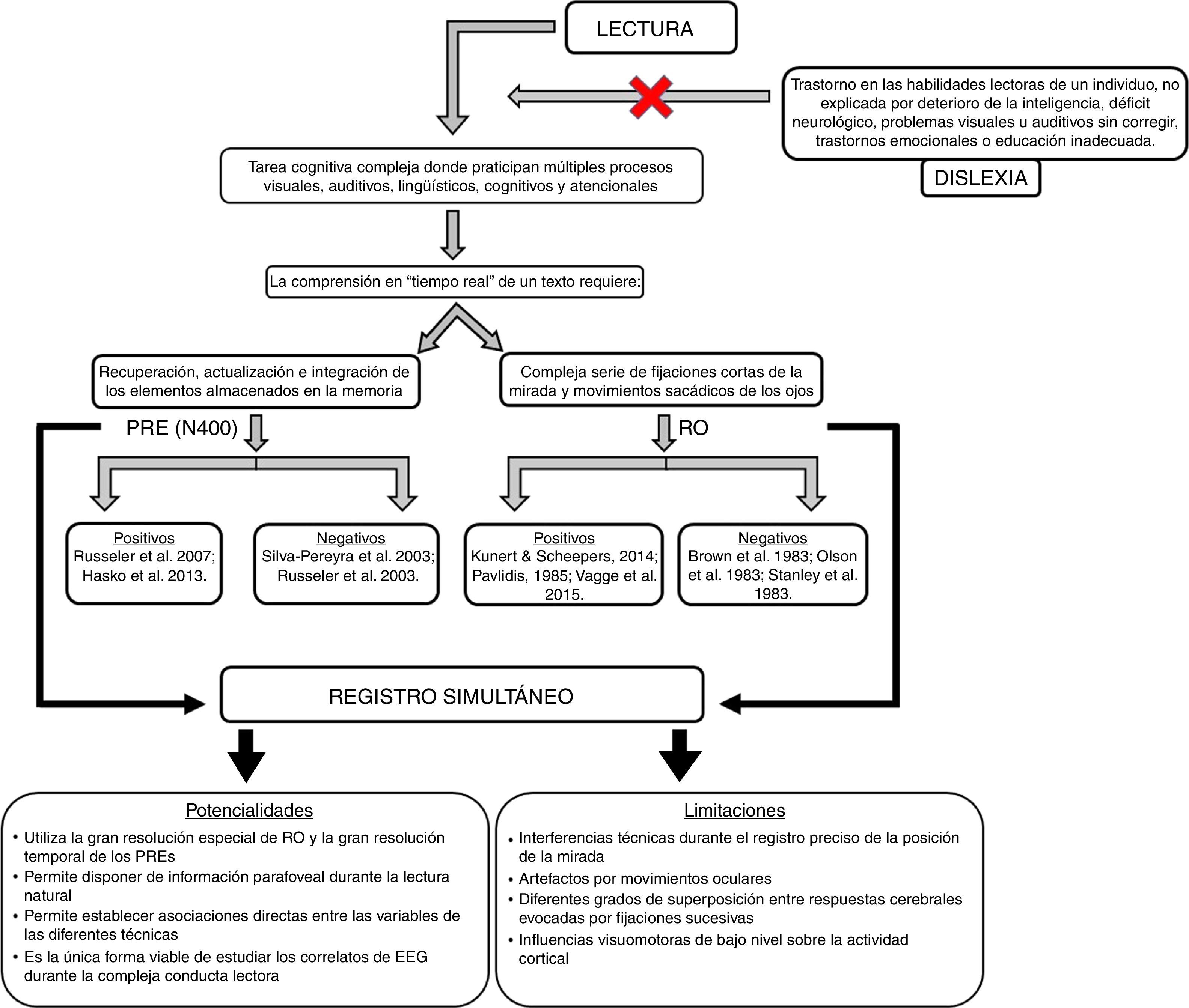

La dislexia es uno de los trastornos del desarrollo más comunes en la etapa escolar y afecta aproximadamente del 5 a 15% de la población. La comprensión en «tiempo real» de un texto requiere del procesamiento de la información visual entrante a través de una compleja serie de fijaciones cortas de la mirada y movimientos sacádicos de los ojos, así como de la recuperación, actualización e integración de los elementos almacenados en la memoria. Esto ha provocado que numerosas investigaciones hayan hecho un gran uso de 2 métodos en particular para su estudio: los potenciales cerebrales relacionados a eventos (PRE) y el rastreo ocular (RO). Sin embargo, los resultados todavía son muy variables y, por tanto, el origen de la dislexia, desde el punto de vista neurobiológico, continúa siendo aún un tema muy debatido. En este trabajo se realiza una revisión exhaustiva de la literatura donde se abordan los hallazgos de los diferentes estudios neurofisiológicos publicados en el tema, así como sus limitaciones. Luego se analiza la factibilidad del registro simultáneo de la actividad eléctrica cerebral y los movimientos oculares mediante la revisión detallada de los aspectos metodológicos a considerar. Por último, se fundamentan las posibles ventajas de este enfoque para los estudios de la lectura, así como sus aplicaciones potenciales en la investigación en dislexia.
Dyslexia is one of the most common developmental disorders at school age and affects approximately 5-15% of the population. Reading is a complex cognitive task and its comprehension requires the processing of visual input across a complex series of brief fixation pauses and saccadic eye movements, as well as retrieving, updating, and integrating contents of memory. This has led to current research making heavy use of two methods: recording eye movement (EMs) and event-related brain potentials (ERPs). However, results are still equivocal, and therefore, the neurobiology of dyslexia remains hotly debated. This paper presents a comprehensive review of the literature in which the findings of different neurophysiological studies published on the subject are addressed, as well as their limitations. The feasibility of simultaneously recording electrical brain activity and eye movements is then analysed by a detailed review of methodological challenges to consider. Finally, the possible benefits of this approach to reading studies and its potential applications in research into dyslexia are described.
Los trastornos en la lectura plantean un alto riesgo para el éxito en el desarrollo individual. Usualmente tienen repercusiones psicosociales, ocupacionales y académicas de largo alcance en los individuos afectados1. A lo largo de los años se ha acumulado un gran número de investigaciones para entender las posibles causas y el alcance de los trastornos de la lectura. La mayor parte de estas investigaciones se han dedicado a la dislexia, la cual se caracteriza por un trastorno en las habilidades lectoras de un individuo, no explicada por deterioro de la inteligencia, déficit neurológico, problemas visuales o auditivos sin corregir, trastornos emocionales o educación inadecuada2-4. La dislexia es uno de los trastornos del desarrollo más comunes en la etapa escolar5,6 y afecta aproximadamente del 5 al 15% de la población7. Además, ocurre en todos los idiomas conocidos8,9 y muestra un patrón típicamente persistente durante toda la vida10.
Actualmente, la etiología de la dislexia es aún un tema muy debatido. Sin embargo, aunque es poco probable que exista un único mecanismo causal para este trastorno, hay un consenso general de que la alteración fundamental es de naturaleza fonológica10-15. Adicionalmente, algunos estudios indican que las alteraciones semánticas también contribuyen a la dificultad lectora en los disléxicos16-19.
La gran mayoría de estudios relacionados con la percepción visual y el procesamiento de palabras durante la lectura se han realizado en ortografías consideradas opacas, en las que la correspondencia entre las letras y sus sonidos es baja. Esto es, algunas letras tienen sonidos distintos en dependencia de la palabra (por ejemplo, del inglés man, make, walk, en donde el grafema a representa 3 fonemas distintos/æ/,/eI/y/¿I/). En comparación, el español es una ortografía considerada como transparente para su lectura, con una alta correspondencia letra-sonido (por ejemplo, casa, paraguas, planta, en donde el grafema a, independientemente de su posición o la palabra en la que se encuentre, siempre representa el fonema/a/). Dado que el aprendizaje de la lectura está influido por las características de la ortografía de la lengua y la relación grafema-fonema, cabría esperar que los mecanismos cerebrales que le subyacen sean distintos entre el inglés y el español. Las dificultades en la adquisición del principio alfabético que involucra la conversión grafema-fonema se han relacionado con una lenta velocidad de denominación20 y con una baja fluidez de la lectura21. La baja velocidad lectora se considera como un rasgo fundamental de la dislexia en el español22-24. En ortografías transparentes para la lectura como el español, especialmente en América Latina, o en el idioma alemán, por ejemplo, se observa sin embargo una mayor dificultad para la escritura, dada la posibilidad de codificar un fonema con más de un grafema (como el caso de/s/, representado por c-s-z), tema poco estudiado como influyente en el proceso lector25,26. Considerando lo anterior, parece razonable pensar que la diferencia en la transparencia de lenguas como el inglés y el español podría determinar variaciones en los sustratos neurales funcionales involucrados en el reconocimiento visual de las palabras en cada lengua.
La lectura es una tarea cognitiva compleja y la habilidad para ello se va aprendiendo lentamente durante la niñez, con la participación de múltiples procesos visuales, auditivos, lingüísticos, cognitivos y atencionales27. Al mismo tiempo, la comprensión en «tiempo real» de un texto requiere del procesamiento de la información visual entrante a través de una compleja serie de fijaciones cortas de la mirada y movimientos sacádicos de los ojos, así como de la recuperación, actualización e integración de los elementos almacenados en la memoria28. Esto ha provocado que las investigaciones actuales hayan hecho un gran uso de 2 métodos en particular: los potenciales cerebrales relacionados a eventos (PRE) y el registro de los movimientos de los ojos o rastreo ocular (RO).
Estudios electrofisiológicos sobre el procesamiento semántico/fonológico en la dislexia. Resultados equívocosCon respecto a los estudios electrofisiológicos, las investigaciones sobre el procesamiento semántico y/o fonológico en pacientes disléxicos son escasas y, además, han arrojado resultados bastante variados11,29-32. Por un lado, un grupo de investigaciones han mostrado anormalidades en el componente N400 en niños y adultos disléxicos comparados con buenos lectores17,18,32,33. Otros autores, sin embargo, no han encontrado diferencias en el mismo componente en pacientes disléxicos con respecto a controles sanos34,35. La N400 es un componente bien establecido que describe una deflexión negativa del PRE, el cual es más prominente alrededor de los 400ms después de la aparición del estímulo en regiones centroparietales36. A pesar de que el componente N400 es particularmente prominente en respuesta a violaciones de tipo semántico, estudios posteriores revelaron que no es exclusivamente un índice de la ocurrencia de una incongruencia semántica, sino más bien una respuesta cerebral típica ante la presentación de estímulos potencialmente cargados de significado en cualquier modalidad de estimulación, incluyendo imágenes37. Sin embargo, su amplitud varía sistemáticamente en función de una variedad de factores, como por ejemplo de la predictibilidad de dicho estímulo en el contexto local en el que es presentado. De este modo, la amplitud de la N400 es inversamente proporcional a la probabilidad de cierre de una palabra en un contexto determinado38. La probabilidad de cierre de una palabra se refiere al porcentaje de individuos que completarían esa oración con una palabra específica cuando les ha sido proporcionada la oración de contexto39. Debido a la sensibilidad que presenta la N400 con respecto al contexto, se piensa que su amplitud refleja la dificultad en la recuperación del conocimiento conceptual almacenado en memoria asociado con una palabra o en su integración dentro del contexto de la oración40. Sin embargo, continúa siendo controversial el hecho de si la N400 refleja la facilitación del acceso a características lexicosemánticas41, un proceso poslexical tardío de integración semántica42,43 o una inhibición semántica44, aunque es posible que múltiples mecanismos contribuyan a la N400. Independientemente de cuál sea su interpretación teórica, la N400 brinda información acerca del curso temporal del procesamiento semántico y su aparición puede ser interpretada como el tiempo máximo necesario para el acceso inicial al significado de la palabra.
El componente N400 también es sensible a las relaciones fonológicas45-47. Numerosos estudios en sujetos sanos han demostrado que las congruencias fonológicas tales como la aliteración (palabras que comparten la porción inicial)47,48 y la rima (palabras que comparten la porción final)49-51 producen una reducción de la N400. Con respecto a los estudios en poblaciones disléxicas, algunos autores han encontrado alteraciones en el componente fonológico de la N400 en adolescentes disléxicos30,31,52-54. Mientras, por otro lado, otros autores no han encontrado diferencias entre disléxicos y buenos lectores29.
De forma general, los estudios mencionados sufren al menos de 3 limitaciones. La primera consiste en que la mayor parte de estas investigaciones se han concentrado solamente en los efectos semánticos o en los efectos fonológicos. Solo unos pocos investigadores se han enfocado en cómo estos 2 tipos de efectos interactúan entre sí55-58. La segunda es que solo unos pocos han utilizado «oraciones» en sus diseños experimentales32,56. Cuando se usan oraciones se tiene la ventaja de contar con un estímulo más natural. Además, en la práctica, la disociación de los efectos semánticos y fonológicos es algo problemático, ya que el contexto de la oración que precede a la palabra «diana» (congruente o incongruente) induce a la misma vez predicciones tanto semánticas como fonológicas. De hecho, se ha demostrado que los procesamientos semánticos y fonológicos interactúan entre sí de una manera compleja56. Por esta razón resultaría útil utilizar en un mismo paradigma oraciones con diferentes grados de incongruencia, tanto semántica como fonológica, ya que esto permitiría explorar los posibles efectos individuales y conjuntos que tienen estos atributos sobre el procesamiento cerebral durante la lectura. La tercera limitación surge del tipo de paradigma utilizado, ya que históricamente los estudios electrofisiológicos para evaluar la lectura se han realizado con el clásico paradigma visual en serie (PVS). Este paradigma permite evitar los artefactos en el electroencefalograma (EEG) por movimientos oculares, ya que los sujetos fijan la mirada en el centro de la pantalla y las oraciones van siendo presentadas palabra a palabra. No hay duda de que el PVS ha sido un método extremadamente exitoso para evaluar el correlato electrofisiológico del reconocimiento de la palabra40. Sin embargo, este paradigma representa una simplificación significativa del proceso normal de lectura. Por ejemplo, durante la lectura natural las palabras no son inspeccionadas de una forma estrictamente consecutiva, sino que son fijadas o saltadas frecuentemente y las regresiones hacia palabras anteriores no son raras. Además, a diferencia del PVS, la lectura natural permite el preprocesamiento de las palabras siguientes en la visión periférica o parafoveal59-61. Adicionalmente, en la mayor parte de los estudios los estímulos presentan una frecuencia de 60-150 palabras por minuto, mientras que durante la lectura natural las frecuencias de 250 palabras por minutos no son raras37. Dada la alta resolución temporal de las variables electrofisiológicas, estos diseños experimentales introducen un sesgo que da lugar a la pregunta de ¿en qué magnitud los resultados obtenidos con el PVS pueden ser aplicados a la lectura natural? Estudios recientes han demostrado que es posible registrar el componente N400 durante la lectura natural de izquierda a derecha en sujetos sanos a pesar de los problemas técnicos que impone este método28,61-67. Sin embargo, no tenemos referencia de la evaluación de este tipo de paradigma en pacientes disléxicos.
Una alternativa para resolver las limitaciones antes descritas sería utilizar un paradigma que cuente con oraciones leídas de forma natural por parte de los sujetos y que, además, utilice de forma simultánea oraciones con diferentes grados de incongruencia semántica y fonológica para evaluar el componente N400. Esto permitiría analizar los efectos semánticos y fonológicos por separado y, al mismo tiempo, evaluar el efecto conjunto de ambos procesamientos, lo que redundaría en una mejor comprensión del proceso lector tanto en sujetos sanos como en pacientes disléxicos. Un elemento adicional importante lo constituye la necesidad de realizar este tipo de investigaciones en poblaciones que tengan como lengua materna al español. Este idioma es hablado por más de 500 millones de personas en el mundo y constituye la segunda lengua del mundo por número de hablantes, además de que cada vez son más las personas que aprenden a hablar español68.
Estudios de la función oculomotora en la dislexia. Resultados incongruentesCon respecto a los estudios sobre los movimientos oculares en pacientes disléxicos, se observa un alto grado de incongruencia entre sus resultados. Por un lado, un grupo de investigadores han encontrado anormalidades en los movimientos oculares en pacientes disléxicos69-72, mientras que, por otro lado, otros estudios no han encontrado diferencias entre disléxicos y sus contrapartes sanas73-75. La cantidad, duración y secuencia de las fijaciones y de las sácadas, entre otras variables, son usadas para hacer inferencias acerca de los procesos cognitivos subyacentes76. La eficiencia de los movimientos oculares se desarrolla en paralelo con el avance en la velocidad y precisión de la lectura. Esta eficiencia se manifiesta como una disminución en la cantidad y duración de las fijaciones, una disminución de las regresiones, así como un aumento en la duración de las sácadas77. Por lo tanto, se esperaría que los pacientes disléxicos mostraran una ineficiencia en los movimientos oculares caracterizada por un aumento en la cantidad y duración de las fijaciones y un aumento en las regresiones asociadas a sácadas más cortas, lo cual, de hecho, ha sido reportado por algunos autores71. Sin embargo, la razón fundamental por la cual se observa tanta inconsistencia entre los resultados de estos estudios se atribuye a la gran diferencia interindividual que existe entre los pacientes con dislexia78. De acuerdo a la clasificación de Boder (1973), en solo algunos individuos se observan alteraciones en el procesamiento visual en dependencia del subtipo de dislexia que presenten79. Los pacientes con dislexia visual o diseidética son los que han mostrado un patrón atípico de movimientos oculares; sin embargo, este subtipo solo se observa en un 10% de los pacientes79,80. Por tal motivo, algunos autores han sugerido que las alteraciones en los movimientos oculares de estos pacientes no obedecen a una disfunción oculomotora, sino que es secundaria a un defecto en el procesamiento visual72,81. Más aun, otros autores han descartado estas alteraciones como causa del trastorno y han sugerido que constituyen más bien un correlato o quizás hasta un biomarcador7.
Registro simultáneo de la actividad eléctrica cerebral y la actividad oculomotora. Una alternativa para la investigación en disléxicosEn la actualidad, el registro simultáneo de diferentes técnicas es un método cada vez más frecuente dentro de las neurociencias. Este enfoque permite utilizar las ventajas de unas técnicas para paliar las debilidades de otras, además de poder establecer relaciones directas entre variables de diferentes técnicas. En el caso particular de la lectura, los PRE brindan una gran resolución para el procesamiento «temporal», mientras que el RO brinda gran resolución para el procesamiento «espacial», ambos necesarios durante la lectura y comprensión de un texto. El registro simultáneo de estas 2 técnicas podría permitir establecer relaciones básicas, por ejemplo, entre la duración de la fijación sobre la palabra «diana» y la amplitud de la N400.
Hasta el momento, sin embargo, han sido pocos los intentos de registrar de forma simultánea los PRE y el RO en situaciones de lectura natural sin restricciones con el objetivo de comparar directamente la actividad cerebral y el comportamiento oculomotor en función de las propiedades de los elementos fijados. Esto probablemente se debe, al menos, a 4 problemas metodológicos asociados a dichos registros: a) es necesario realizar un registro preciso de la posición de la mirada sin interferencias técnicas, b) artefactos por movimientos oculares, c) diferentes grados de superposición entre respuestas cerebrales evocadas por fijaciones sucesivas, y d) influencias visomotoras de bajo nivel sobre la actividad cortical.
Registro preciso de la posición de la mirada sin interferencias técnicasContar con una información exacta acerca de la latencia y localización de cada fijación constituye un requerimiento básico para las promediaciones basadas en las fijaciones. Tradicionalmente, los investigadores han usado electrodos electrooculográficos (EOG) cerca de los ojos para controlar la fijación estable de la mirada82. Sin embargo, su resolución espacial de 1,5°-2° no es suficiente para ofrecer la posición exacta de la mirada necesaria durante el análisis de la lectura83-85. No obstante, los equipos actuales de rastreo visual cuentan con una resolución espacial de hasta 0,01° con frecuencias de muestreo de hasta 2kHz que sí les permiten ofrecer la exactitud necesaria para estos tipos de análisis86-88.
Otros elementos técnicos a considerar con respecto a este primer problema son: a) los artefactos por presión debido al contacto de los electrodos anteriores con el equipo de rastreo ocular, específicamente en aquellos equipos fijados a la cabeza, b) artefactos musculares por estabilización de la cabeza o posiciones incómodas del sujeto durante el registro, c) una sincronización adecuada entre los sistemas de registros PRE-RO, y d) artefactos electromagnéticos por la cercanía de un equipo eléctrico funcionando cerca de los electrodos del EEG. Estas interferencias pueden ser minimizadas con: a) el uso de un dispositivo acolchonado sobre los electrodos anteriores, b) el ajuste cuidadoso de una posición cómoda de sentado del sujeto, c) la sincronización de los sistemas mediante un pulso denominado en inglés transistor-transistor logic (TTL), compartido cada pocos segundos, y d) la aplicación de un filtro notch en la banda de frecuencia del artefacto89.
Artefactos por movimientos ocularesEstos tipos de artefactos en el EEG son generados por 3 mecanismos: a) la rotación del dipolo corneorretinal del globo ocular90; b) el parpadeo y las sácadas ascendentes91, y c) la actividad eléctrica de los músculos intrínsecos del ojo al principio de la sácada, la cual se observa en el EEG como un potencial de espiga92. Ya que la exclusión de las épocas contaminadas no es una opción durante la lectura natural, se ha propuesto una gran variedad de algoritmos para corregir matemáticamente los artefactos oculares93-97. Más aun, algunos autores sugieren que el registro de rastreo ocular puede ayudar a evaluar, comparar y mejorar los métodos de corrección por al menos 2 razones28. La primera de ellas es que, contrario al EOG, el rastreo visual ofrece una medida de la posición de los ojos que es eléctricamente independiente del EEG. Por lo tanto, las correlaciones entre el rastreo ocular y el EEG ya corregido, probablemente resulten de la actividad artefactual residual tanto miogénica como corneorretinal. Es decir, el grado en el que el EEG (después de la corrección) depende de la posición de los ojos, puede ayudar al investigador a evaluar la calidad de la corrección entre los estudios y los algoritmos. Y la segunda razón radica en el hecho de que el sistema de rastreo visual puede ofrecer información acerca de si un algoritmo sobrecorrigió los datos o distorsionó la verdadera actividad cerebral. Debido a que un experimento dado posee, al menos, algunos intervalos con una fijación estable de la mirada, los sistemas de rastreo visual de alta resolución le permiten al investigador seleccionar de forma objetiva intervalos del EEG que estén libres de artefactos oculares (parpadeos, sácadas y microsácadas). Estos intervalos no deben ser alterados por la corrección ocular y, por lo tanto, ofrecen una medida para cuantificar la distorsión de las señales cerebrales introducidas por el método.
Superposición entre respuestas cerebrales evocadas por fijaciones sucesivasLa presentación en serie de las palabras (PVS) ofrece un control total sobre el estímulo que está siendo presentado al sistema visual en un momento dado. Sin embargo, durante la lectura natural el experimentador tiene muy poco control sobre el patrón espaciotemporal de las fijaciones, en particular sobre la latencia y duración a la que un sujeto elige fijar una palabra diana. Esto conlleva a 2 problemas de superposición del EEG:
El primer problema es el grado de superposición temporal entre el potencial evocado por la fijación a una palabra específica y la actividad electroencefalográfica de base que está relacionada, por ejemplo, a la aparición de la oración. El segundo problema es la superposición temporal entre los potenciales evocados por las fijaciones sucesivas a distintas palabras en la oración. En el primer caso, algunos autores han observado que la aparición de la oración evoca una P300 que solo regresa a nivel de la línea de base alrededor de los 700ms. Por lo tanto, pudiera ocurrir que las palabras dianas en 2 condiciones diferentes sean fijadas a latencias sistemáticamente distintas después de la aparición de la oración, como por ejemplo en el caso de oraciones con cantidad de palabras distintas o con diferentes localizaciones de la palabra diana dentro de la oración. Es decir, los potenciales relacionados con esas fijaciones ocurrirían sobre actividades de base diferentes, lo que distorsionaría la forma de onda y la topografía entre diferentes condiciones. Con respecto al segundo caso, un intervalo entre fijaciones de alrededor de 250ms (como normalmente ocurre durante la lectura natural) conlleva a una superposición masiva de los potenciales provocados por fijaciones sucesivas. Esto significa que los componentes tardíos evocados por la fijación a la palabra anterior a la diana (n-1) se van a superponer a los componentes tempranos evocados por la fijación a la palabra diana (n); de igual manera, los componentes tardíos evocados por la fijación a la palabra diana se van a superponer a los componentes tempranos evocados por la fijación a la palabra posterior a la diana (n+1).
A pesar de que han sido propuestos numerosos métodos de deconvolución para separar los potenciales superpuestos en experimentos de PRE con estimulación rápida e intervalos interestímulos variables98-101, hasta la fecha no existe una solución simple a este problema de la superposición, el cual incluso ha sido ignorado en estudios previos. Sin embargo, también hay que tener en cuenta que la magnitud en la que la superposición se convierte en un problema depende al menos del tipo de paradigma usado. Por ejemplo, en el tema que nos atañe, una alternativa útil seria utilizar un paradigma en el cual sus diferentes condiciones sean lo más similares posible entre ellas, es decir, oraciones con igual cantidad de palabras, con localización de la palabra diana al final de la oración y con las palabras dianas con igual número de sílabas.
Influencias visomotoras de bajo nivel sobre la actividad corticalLos potenciales evocados relacionados con las fijaciones y las sácadas no son modulados solamente por las demandas del procesamiento cognitivo superior. La forma de onda de dichos potenciales también es influenciada por factores visomotores de bajo nivel que no pueden ser controlados durante el comportamiento natural de escaneo visual. Esta influencia debe ser cuidadosamente delineada de aquellos efectos que realmente reflejen un procesamiento cognitivo superior (por ej., procesamiento semántico) de las palabras específicas durante la lectura. Dentro de estos factores de bajo nivel se encuentran: las propiedades exactas de la información visual entrante durante la sácada y la cinemática de la sácada.
Con respecto al primero, se conoce que en la mayor parte de los estudios de PRE que evalúan el procesamiento cognitivo superior, se le presta mucha atención al emparejamiento de los estímulos visuales en términos de propiedades de bajo nivel como la luminosidad, contraste y frecuencia espacial. Por ejemplo, tanto la amplitud como la latencia de la respuesta visual lambda, que ocurre aproximadamente a los 100ms luego del inicio de una fijación, varían en función de la luminosidad, contraste y frecuencia espacial del fondo102-104. Hasta donde se conoce, no ha sido investigado si las diferencias en la información visual que alcanza la fóvea durante las fijaciones en diferentes localizaciones (por ej., cambios locales en la luminosidad y contraste por mirar diferentes partes de una escena) causan modulaciones relevantes sobre los potenciales cerebrales. Sin embargo, en el caso particular del registro de los PRE durante la lectura natural, se puede asumir que las propiedades del campo visual sean comparables para las diferentes palabras dentro de la pantalla. Adicionalmente, se pueden intentar reducir los efectos puramente visuales mediante el uso de una máscara que cubra el campo visual periférico.El segundo factor importante de bajo nivel que influencia los potenciales es el tamaño de la sácada. La respuesta lambda, en particular, comprende tanto una respuesta relacionada con el inicio de la sácada, como otra relacionada con su desplazamiento105,106. Durante grandes sácadas (>10°), esta respuesta se disocia en 2 subcomponentes. El primero, presumiblemente relacionado con cambios visuales al inicio de la sácada, y el segundo, de mayor tamaño, presumiblemente evocado por la entrada de nueva información visual durante el desplazamiento del globo ocular106. Por lo tanto, hay que ser muy cuidadoso cuando se comparan condiciones que difieran en términos de amplitud de la sácada. Sin embargo, esto no necesariamente constituye una limitación fundamental para los experimentos de lectura, ya que durante la misma, las sácadas tienen una amplitud bastante constante de aproximadamente 7 a 9 caracteres76.
A pesar de estos retos metodológicos, recientemente un grupo de investigaciones han demostrado la factibilidad de realizar este corregistro PRE/RO durante la lectura natural de oraciones de izquierda a derecha28,62-67. Incluso han demostrado la ventaja que supone este tipo de corregistro, ya que permite investigar aspectos de la lectura que sería difícil o imposible de hacer con el clásico PVS. Un ejemplo de esto, como se describió anteriormente, es la disponibilidad de la información parafoveal durante la lectura natural. La duración y la magnitud a la cual son preprocesadas las palabras que aún no se han leído constituye todavía un tema controversial, que a la vez puede ser estudiado con este tipo de corregistro. Es decir, la pregunta de si el significado de la palabra puede ser extraído a partir de las palabras parafoveales es un tema que puede ser investigado con este enfoque y, al mismo tiempo, puede ser importante en trastornos como la dislexia. Además, el registro simultáneo es la única forma viable de estudiar los correlatos del EEG durante la compleja conducta lectora. Por ejemplo, preguntas interesantes como: ¿cuál es el correlato electroencefalográfico de una palabra cuando es saltada, en vez de cuando es fijada?, ¿cuáles son los potenciales relacionados con la fijación que preceden o siguen a la decisión de desencadenar una sácada regresiva? o ¿cuál es la localización funcional de las diferencias individuales en la habilidad y velocidad durante la lectura?, solo pueden ser abordadas con este enfoque (tabla 1).
Principales hallazgos de la presente investigación
| El origen de la dislexia, desde el punto de vista neurobiológico, continúa siendo un tema muy debatido |
| La mayor parte de las investigaciones publicadas sobre la dislexia no se han realizado en idiomas considerados como transparentes, por ejemplo, el español |
| Los métodos de registro de la actividad eléctrica cerebral y de los movimientos oculares han demostrado gran utilidad en la demostración objetiva de las alteraciones en disléxicos; sin embargo, los resultados son muy variables aún |
| Una limitación importante en el caso de los estudios electrofisiológicos es que generalmente no se realizan en condiciones de lectura natural |
| El corregistro de la actividad electroencefalográfica y la actividad oculomotora puede constituir una alternativa para la investigación en dislexia |
| A pesar de que se ha demostrado que el método de corregistro es factible, hay que considerar las limitaciones metodológicas que este enfoque impone |
Sin embargo, a pesar de que estos estudios realizaron una descripción detallada de las nuevas posibilidades que se abren con este método de registro simultáneo, hasta el momento no existen reportes de que se haya utilizado en poblaciones clínicas. La dislexia, en particular, es un trastorno que se puede beneficiar mucho de esta nueva alternativa siempre y cuando se realice bajo un paradigma que permita responder a las preguntas que todavía no tienen respuestas concretas. Adicionalmente, tampoco existen reportes de que este método se haya utilizado en el idioma español, por lo que sería una alternativa interesante, especialmente considerando que la dislexia no se comporta igual entre los diferentes idiomas107(fig. 1).
Conclusiones
La percepción visual fuera del laboratorio, y en particular la lectura, es fundamentalmente transacádica y comprende un muestreo activo, dinámico y rápido de la escena visual. Qué tipos de informaciones pueden ser obtenidas a partir de cada fijación y cómo estas informaciones pueden ser integradas a los datos de fijaciones previas y subsiguientes, aún constituyen preguntas de difícil explicación, pero pueden ser abordadas mediante el registro simultáneo de la actividad electroencefalográfica y la actividad oculomotora. Sin embargo, a pesar de las ventajas que supone este enfoque, hay que tener mucho cuidado a lo hora de diseñar metodológicamente un experimento debido a las dificultades técnicas que impone este tipo de corregistro. La dislexia es un trastorno que cada vez cuenta con mayor atención dentro de la comunidad científica debido a su impacto en diferentes esferas de la vida, pero que continúa teniendo un vacío con respecto a las bases neurobiológicas de su origen. Por tal motivo, con las consideraciones apropiadas tanto desde el punto de vista técnico como analítico, esta alternativa podría contribuir con respuestas nuevas a preguntas viejas.
Conflicto de interesesLos autores declaran que no existen conflictos de intereses.