


Las infecciones producidas por virus se encuentran entre aquellas más difíciles de tratar y en las que sólo muy recientemente se está obteniendo un cierto éxito. Con seguridad, los próximos años serán muy fructíferos en este campo y por eso resulta interesante hacer una descripción de los fármacos antivirales de que se dispone en el umbral del nuevo siglo y los caminos que se vislumbran. Éstos se tratan agrupados por su capacidad de interacción sobre la/s membrana/s o el genoma viral,
con una organización en función de su estructura molecular, que se relaciona con el mecanismo por el que actúan.
Los virus son ácidos nucleicos rodeados por una cubierta proteica, son entonces «paquetes de información genética», que se introducen en una célula huésped y dirigen la maquinaria metabólica de esta para producir más virus, siendo por ello parásitos obligados. En el medio extracelular los virus son inactivos y presentan la morfología característica del grupo al que pertenecen, en este estado se habla de partícula viral o virión.
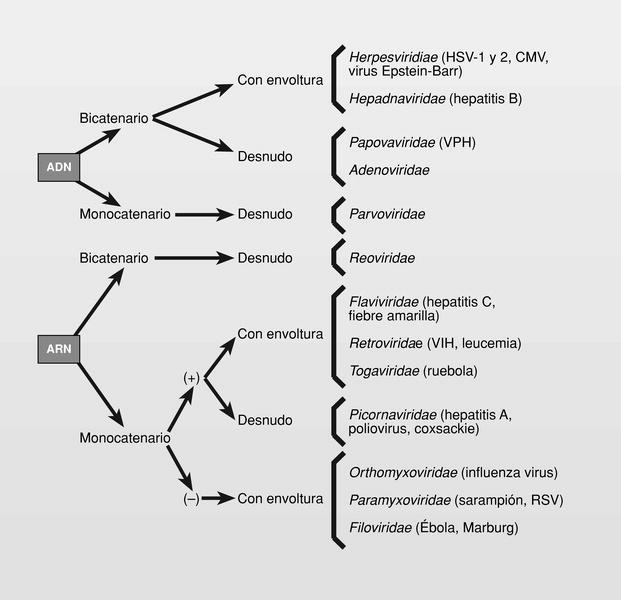
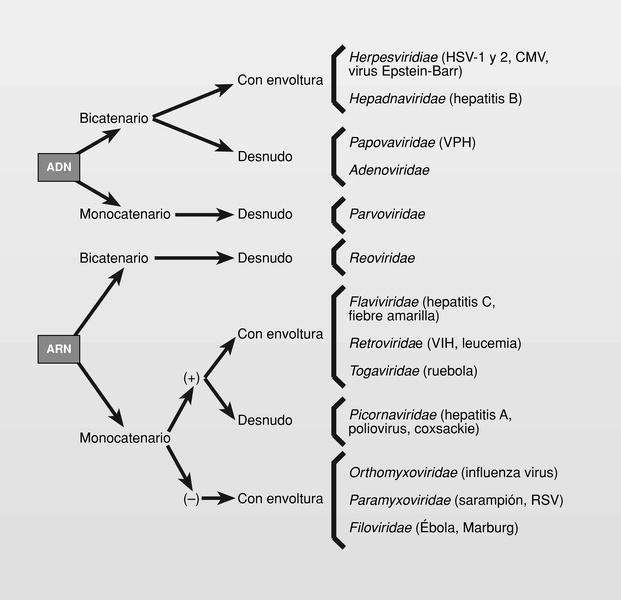
La estructura básica de un virión es un ácido nucleico --puede ser ADN o ARN (nunca ambos) y bicatenal o monocatenal-- rodeado por una proteína (cápside) y a veces por una membrana (envoltura) en cuya composición participan lípidos, hidratos de carbono y proteínas. Los virus que carecen de esta envoltura, se llaman «virus desnudos»1. Estos sencillos aspectos comentados nos hacen ver la simplicidad, pero también la variabilidad con que podemos encontrarnos. En este trabajo se describe la quimioterapia antiviral disponible para virus capaces de producir enfermedades en humanos2, nombrados siguiendo las normas establecidas por el Comité Internacional de Taxonomía de los Virus (ICTV) creado en 1966 (fig. 1) y no tratará de vacunas3. Los mayores problemas con que nos encontramos al tratar de establecer una quimioterapia antiviral son:
Los virus, al tener una constitución tan simple, no ofrecen grandes posibilidades de puntos de ataque.
Al tener que producirse su proliferación a expensas de la célula huésped, es difícil establecer una acción específica.
Al producirse la primera sintomatología cuando ya se superó un punto límite, los fármacos al uso son menos efectivos, y aquellos más interesantes serían los preventivos.
A todo esto hay que añadir que hasta mediados de los años cincuenta no era posible el ensayo de nuevas moléculas en sistemas de cultivo celular4. Así, podemos x decir que fue en las décadas de los sesenta y setenta cuando aparecieron los primeros compuestos activos, pero realmente el desarrollo de este tipo de quimioterapia tiene lugar en la última década.
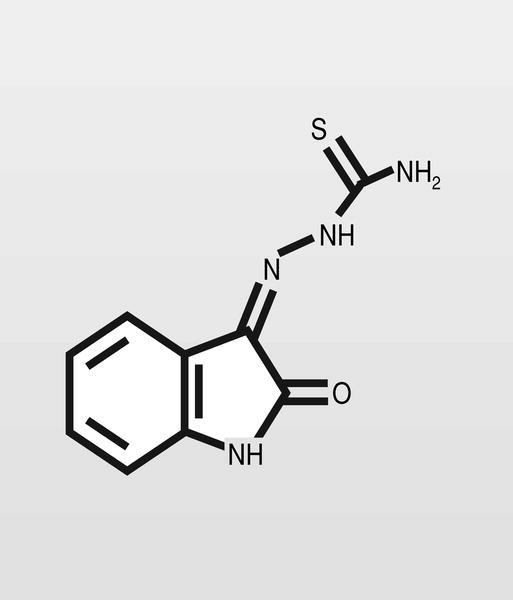
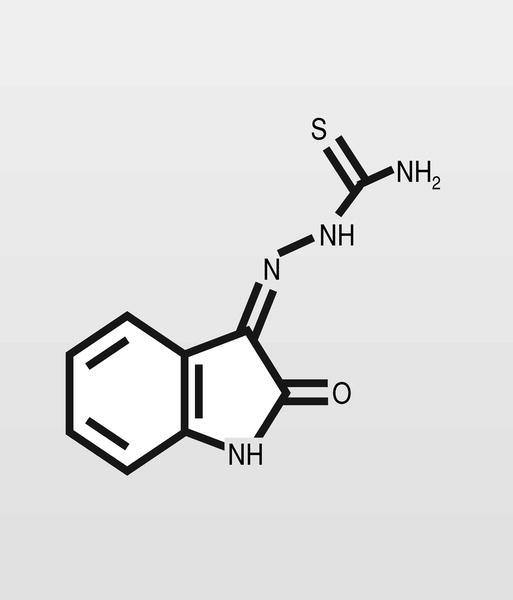
Las primeras moléculas descritas con actividad antiviral fueron las tiosemicarbazonas, que eran estudiadas por su actividad antimicrobiana a finales de los años cincuen ta. La metisazona (fig. 2) fue el primer antiviral ensayado extensamente en humanos, se hizo al inicio de la década de los sesenta en la India y frente a ADN-virus. En cualquier caso, de estas primeras moléculas se mantienen prácticamente sólo los análogos de nucleósidos, que son ampliamente utilizados en la actualidad y que se describirán con detalle.
Para tratar de organizar el estudio del conjunto de los fármacos antivirales disponibles, considero oportuno establecer una clasificación básica que los diferencie por el modo de cómo afectan a los dos componentes esenciales de los virus, y utilizar después un criterio de estructura, que obviamente definirá su mecanismo de acción:
Fármacos que interfieren sobre componentes de membrana. Fundamentalmente influirán sobre las fases de introducción --adsorción, penetración y descapsidación-- y también sobre las fases de maduración y liberación.
Fármacos que interfieren sobre el genoma viral. Fundamentalmente influirán sobre los mecanismos de replicación.
Fármacos que intervienen sobre componentes de membrana
Los componentes de membrana van a tener importancia, en primer lugar porque van a ser los sistemas de reconocimiento del virus por la célula huésped y los que permitan su entrada en ella, y también porque finalmente va a ser necesario reconstruir esta membrana, en el proceso de maduración, para generar el virión que ha de liberarse. En muchas de estas fases se encuentran dianas terapéuticas, y muchas de ellas están en desarrollo, pero se comentan casi exclusivamente aquellas para las que actualmente se dispone de fármacos de uso clínico.
Inhibidores de la adsorción
En el caso del virus de la inmunodeficiencia humana (VIH), por ejemplo, la adsorción se realiza porque se produce un reconocimiento gp120 del virus-receptores CD4 de la membrana de los linfocitos T, de modo que una estrategia terapéutica fue dirigida a impedir ese proceso:
Análogos de CD4. Aunque se dispone de buenos, presentan problemas importantes porque los receptores CD4 no son solo receptores para los virus.
Inhibidores del complejo de interacción gp120-CD4. Se conocen diversas moléculas, que además, como no hacen el bloqueo de manera demasiado específica, tienen actividad también frente a otros tipos de virus con envoltura:
Polisulfatos de polisacáridos.
Polisulfonatos. Un prototipo es la suramina, que fue la primera molécula reconocida como anti-VIH y el primer fármaco usado para el tratamiento del sida5, aunque pensando erróneamente que era inhibidor de transcriptasa inversa (TI).
Ciertos complejos metálicos.
Inhibidores de la penetración-descapsidación
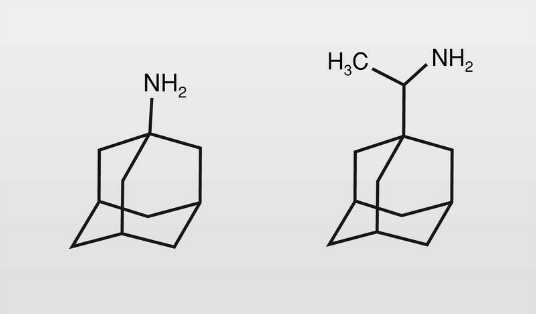
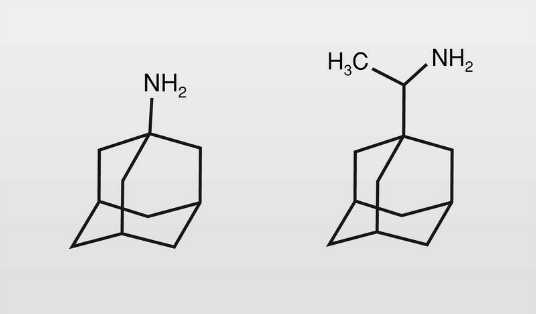
Los derivados del adamantano (fig. 3) son fármacos de este tipo porque impiden la entrada del virus en la célula huésped al bloquear la descapsidación del virus. Tienen utilidad contra el virus de la gripe:
La hemaglutinina viral «reconoce» al ácido siálico presente en la membrana de la célula que va a ser atacada y el virus es introducido. Posteriormente se forma un canal iónico (M2) en la cubierta viral, que permite la entrada de protones y cuando el pH baja, se inducen cambios --hidrólisis ácida-- en la hemaglutinina, que permite la liberación del genoma viral. Estos compuestos, bloquean el canal M2, impidiendo así el desnudamiento viral. A concentraciones más altas, hay también procesos de interferencia en la liberación.
La amantadina (1-aminoadamantano) se introdujo en 1966 y no fue muy utilizada debido a su limitado efecto, a los problemas sobre el SNC (mareos, insomnio) y el SNA --tiene también un limitado uso como antiparkinsoniano por su efecto anticolinérgico-- y a la aparición de resistencias.
La rimantadina se introdujo en clínica en 1994, siendo su dosis necesaria aproximadamente la mitad que de amantadina y no presenta tantos problemas secundarios.
Dentro de los inhibidores de la penetración, podemos incluir también como fármacos de reciente uso clínico para el tratamiento del virus de la gripe, los inhibidores de neuraminidasa6 aunque también podrían ser considerados, como se hace en algunas monografias, fármacos inhibidores de la liberación.
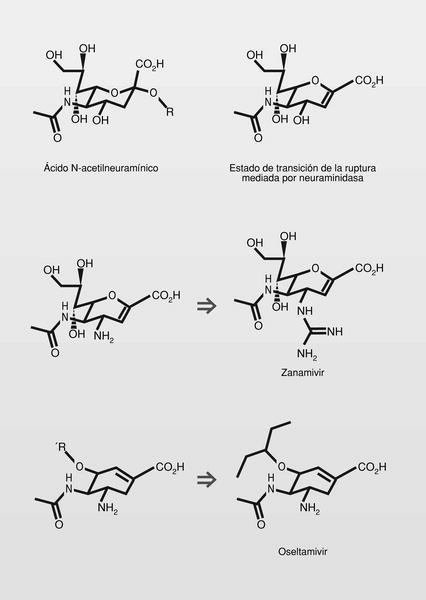
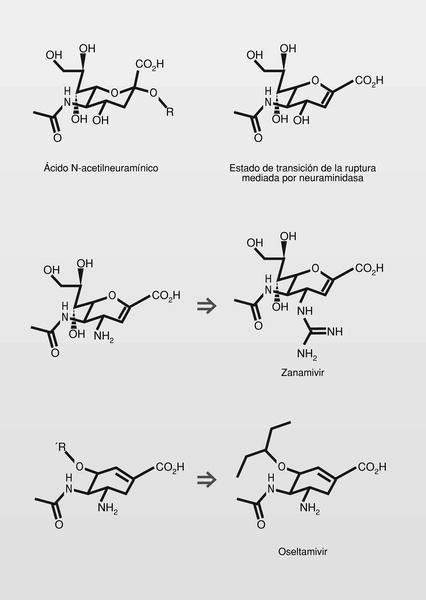
Los glicoconjugados (glicoproteínas y glicolípidos) son elementos de membrana de todas las células de vertebrados y actúan como ligandos de reconocimiento de proteínas sean internas o externas (por ejemplo, para el reconocimiento de patógenos). Estos glicoconjugados en células de mamíferos terminan generalmente por ácidos siálicos, entre los que el más importante es el ácido N-acetilneuramínico. El virus de la influenza infecta células porque se une a esos ácidos siálicos de superficie utilizando una proteína viral de enlace, la hemaglutinina. El virus necesita también una «enzima destructora de receptor», en este caso la neuraminidasa, que hidroliza esos ácidos siálicos terminales y permite su entrada en la célula. Esta neuraminidasa viral es necesaria también para la liberación del virus una vez formado en el interior de la célula.
Ambas proteínas virales en el caso del virus de la influenza (hemoglutinina y neuraminidasa) fueron determinadas por rayos X en su interacción con sus receptores y ello las convierte --especialmente a la neuraminidasa-- en elementos muy útiles para el diseño racional de inhibidores que puedan dar lugar a fármacos antivirales
La sustitución del hidroxilo de la posición 4 del ácido N-acetilneuramínico, por un grupo amino, incrementó mucho la potencia inhibidora de la neuraminidasa (aproximadamente 200 veces), y la sustitución por un grupo guanidino (aproximadamente 5.000 veces). Además, el zanamivir resultó un millón de veces más potente en su unión a la neuraminidasa del virus que el propio ácido N-acetilneuramínico. Su hidrofilia dificulta su absorción por vía oral y por ello se administra por vía respiratoria.
Como se observa en la figura 4, en el estado de transición de la ruptura del ácido N-acetilneuramínico por la enzima, el ciclo es casi planar en el entorno del átomo de oxígeno, como resultado de la tensión originada en el ciclo de azúcar durante ese estado de transición (a ello contribuyen también las fuertes interacciones iónicas entre el grupo carboxilo y tres residuos de arginina del sitio de reconocimiento y el doble enlace formado). Esta conformación se mimetiza bien con un anillo de ciclohexeno, en el que el doble enlace se sitúe hacia la posición que ocupaba en el ciclo el átomo de oxígeno. Derivados del ciclohexeno son estructuras más estables e incluso más versátiles desde el punto de vista químico y sintético.
En uso clínico muy reciente se encuentra también el oseltamivir (fig. 4). El grupo pentiloxi de la posicion 3, además de aumentar su lipofilia y hacer el compuesto absorbible por vía oral, se aloja en un área hidrofóbica del receptor, por lo que mejora sus características de enlace6.
Fármacos que impiden la maduración
Son fármacos que actúan después de que todos los componentes del virión hayan sido ya sintetizados por la célula huésped, pero que consiguen impedir que se ensamblen adecuadamente.
Inhibidores de la proteasa del VIH5b
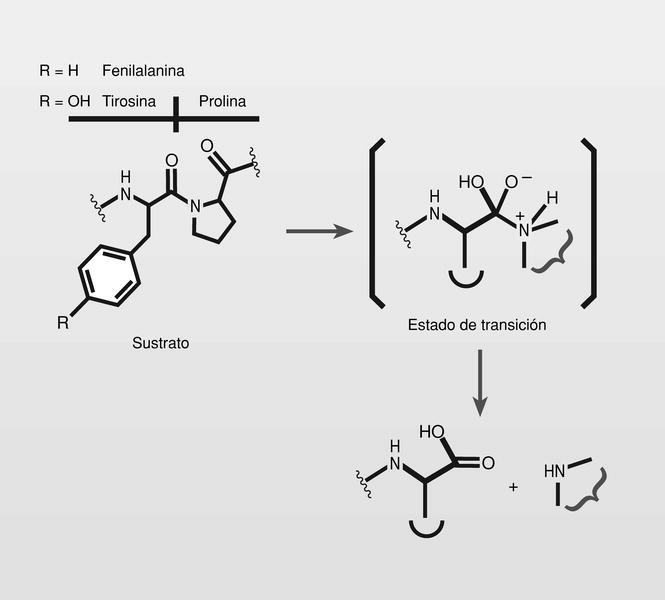
Una aspartil-proteasa codificada por un gen viral pol (el gen pol codifica enzimas: proteasa, integrasa, ribonucleasa, TI) es la responsable de la ruptura de proteínas precursoras codificadas por el gen viral gag (gag codifica las proteínas de estructura) para dar lugar a las proteínas maduras, y su inhibición podría causar grave daño viral. Esta es una estrategia que se está desarrollando extensivamente porque estas proteínas son del tipo de otras aspartil-proteasas humanas ya conocidas (renina).
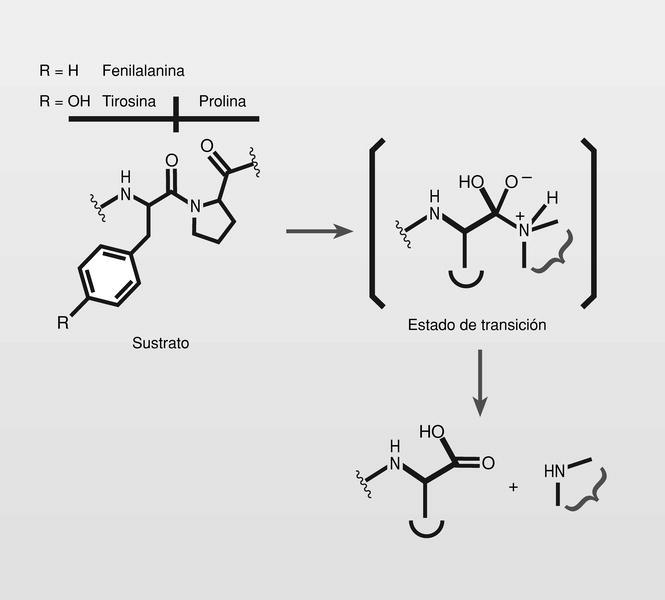
Por otra parte, los puntos de ruptura de esta proteasa viral fueron identificados, presentando alta especificidad por las uniones [Tyr-Pro] y [Phe-Pro], lo que fue usado para el diseño de inhibidores apropiados. Para ello fue seguido el principio de «peptidomiméticos del estado de transición» y tratando de reemplazar la unión peptídica hidrolizable por un isóstero no hidrolizable de ese estado de transición (fig. 5).
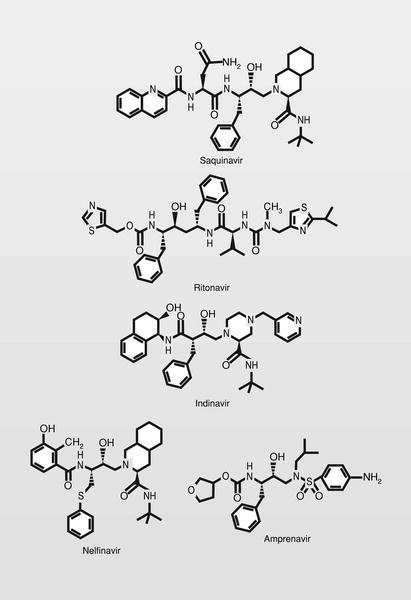
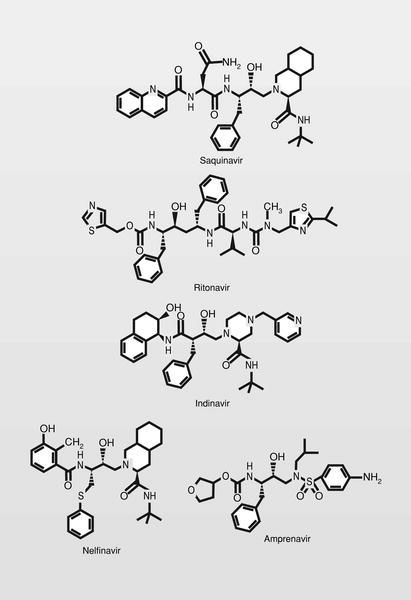
Fueron sintetizados y estudiados varios inhibidores de la proteasa del VIH conteniendo un isóstero del estado de transición hidroxietilamínico y varios de ellos son de uso clínico en la actualidad (fig. 6), normalmente en terapia combinada con los más clásicos análogos de nucleósidos.
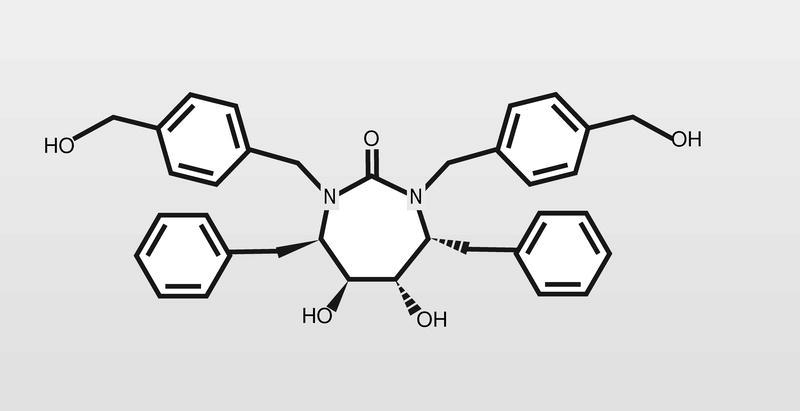
Este tipo de fármacos de tipo peptídico presenta generalmente una corta vida media (debido a la acción de enzimas proteolíticas) y pobre absorción por vía oral. En muchos casos tratan de «enmascararse» (eliminando grupos hidroxilos, incluyendo grupos lipófilos u obteniendo profármacos) cuando ello es posible. De todas formas, se están haciendo esfuerzos diferentes a esta aproximación peptídica, y una de las más importantes es la de ureas cíclicas no peptídicas, entre las cuales podemos considerar al XM323 (fig. 7) como un cabeza de serie y fármaco en fase clínica de estudio con buena absorción por vía oral.
Fármacos que impiden la miristoilación
Las proteínas virales necesitan ser finalmente miristoiladas para que los viriones sean maduros. El ácido mirístico se une al N de una glicina terminal mediante una N-miristoiltransferasa. Se conocen varios inhibidores de la miristoilación pero todos a tan alta concentración que no tienen utilidad terapéutica.
Fármacos que impiden la glicosilación
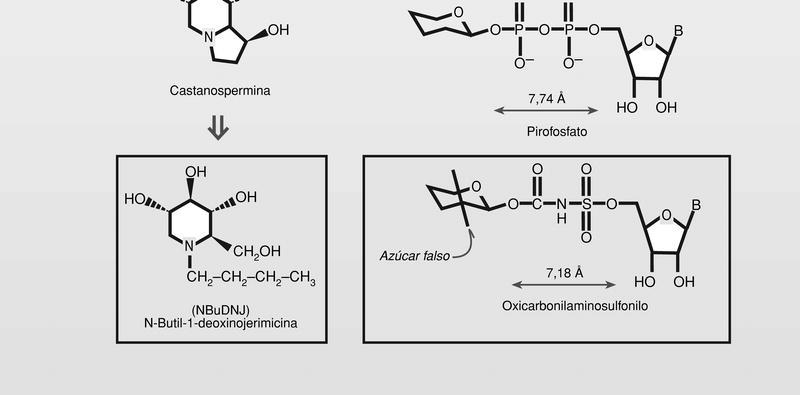
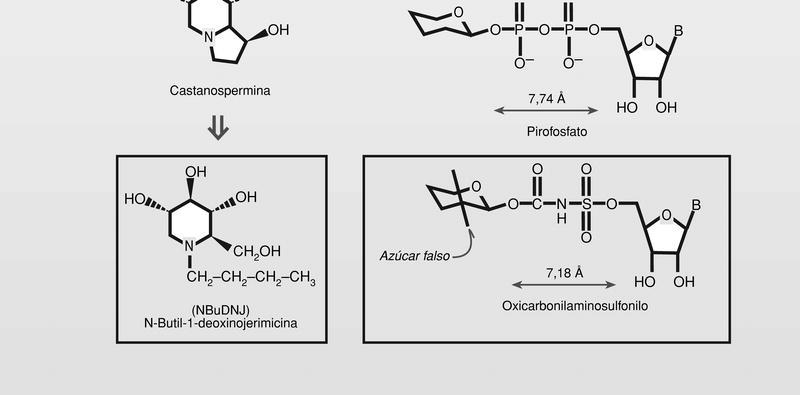
La glicosilación necesaria para la formación de las glicoproteínas de la cápside y de la envoltura, en su caso, es un proceso que también ha sido tomado como diana de la intervención farmacológica, aunque hasta el momento no ha dado resultados terapéuticos. Se trata en general de aminoazúcares, que se comportan como azúcares falsos, que además, pueden introducirse en análogos de nucleósidos difosfatados de unas características estéricas parecidas y menor polaridad que los transportadores normales, por lo que estaría facilitado su paso al citoplasma celular (fig. 8).
Fármacos que intervienen sobre el genoma viral
El grupo de fármacos antivirales que actúan interfiriendo sobre el genoma viral es en la actualidad el más numeroso, y también en el que hay una diferencia más clara entre los distintos tipos de virus, especialmente los retrovirus como el VIH, y los demás. Específicamente existe en los retrovirus la transcriptasa inversa, que es diana del más importante grupo de fármacos anti-VIH.
En general son antimetabolitos, es decir, análogos de metabolitos naturales, que por similitud estructural los sustituyen e impiden que cumplan su función.
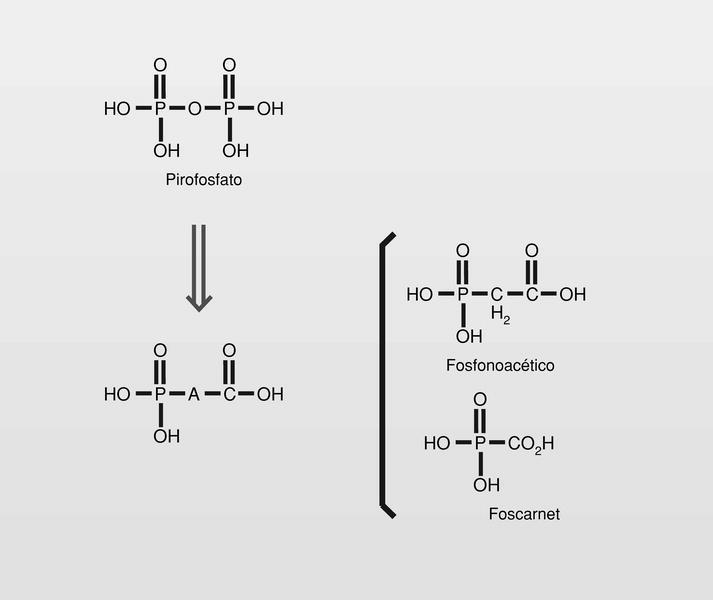
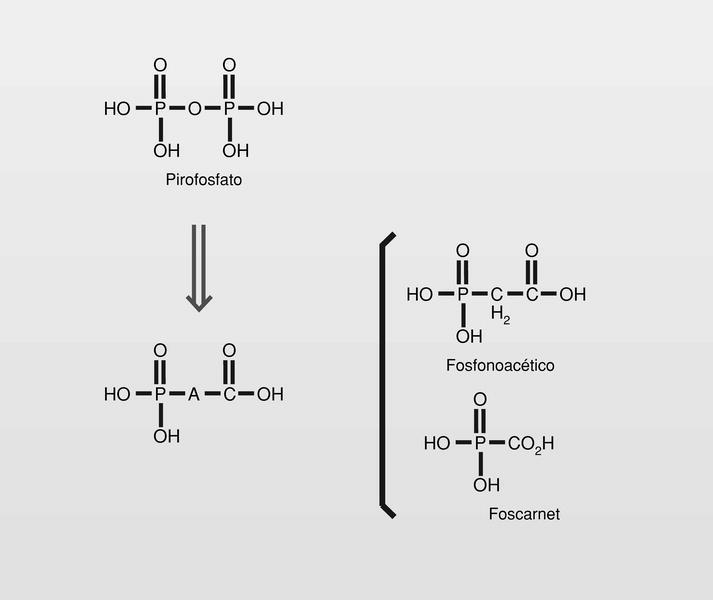
Análogos de pirofosfato
Moléculas que pueden ocupar el sitio de unión del pirofosfato en el ADN o ARN-polimerasa, impidiendo por ello su función (fig. 9).
El foscarnet (sal trisódica del ácido fosfonofórmico) (fig. 9) inhibe la polimerasa viral 100 veces más que la polimerasa celular. Cuando el trifosfato de nucleótido se une a la polimerasa, el fragmento pirofosfato se rompe y el monofosfato se une a la cadena de ADN o al ARN que está en fase de polimerización. Si sobre la polimerasa no hay sitio para aceptar el pirofosfato porque ese sitio está ocupado por el fármaco, no se produce la hidrólisis del pirofosfato y se bloquea la polimerización.
Análogos de nucleósidos
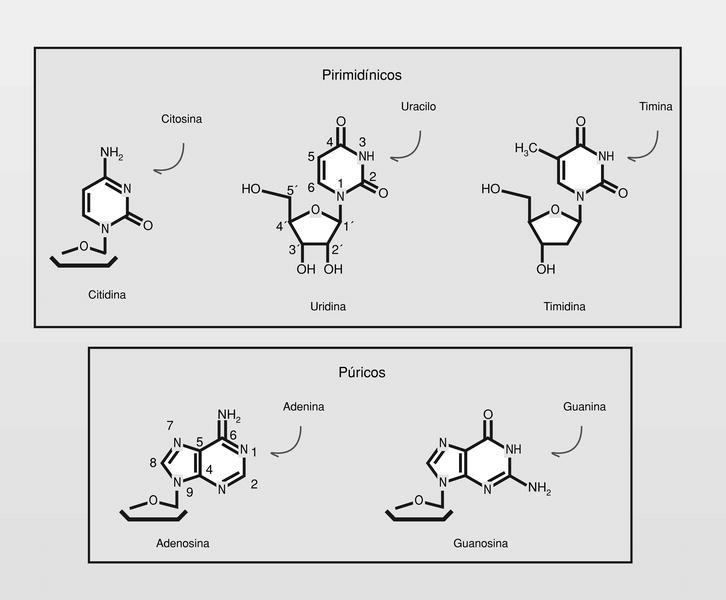
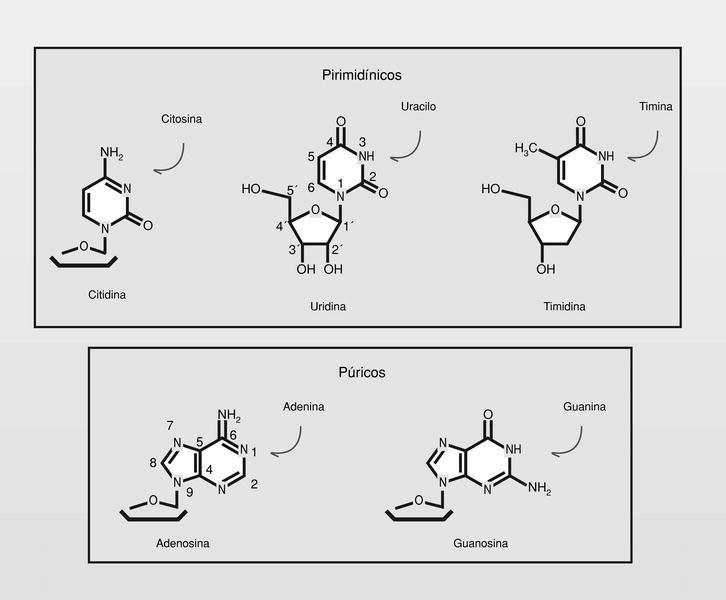
Los nucleósidos son componentes de los ácidos nucleicos (y también de otras macromoléculas bioquímicas), compuestos por un azúcar, ribosa o desoxiribosa según sea ARN o ADN, respectivamente, y una base heterocíclica púrica --adenina, guanina-- o pirimidínica --citosina, uridina o timina-- según se trate de ARN o ADN (fig. 10).
Es entonces fácil imaginar cómo análogos de estos nucleósidos pueden comportarse cómo antimetabolitos y modificar o anular las funciones de los nucleósidos naturales, interfiriendo con los procesos de traducción y/o transcripción en los que están implicados.
Las modificaciones estructurales que pueden producirse son esquemáticamente de tres tipos: sobre la base, sobre el azúcar y sobre el modo de unión entre la base y el azúcar (fig. 10). De los miles de nucleósidos sintetizados, solo unos pocos están utilizándose actualmente en clínica (aunque muchos otros presentan también interesante actividad antiviral --también anticáncer-- in vitro y/o in vivo), y es a los que se hará referencia.
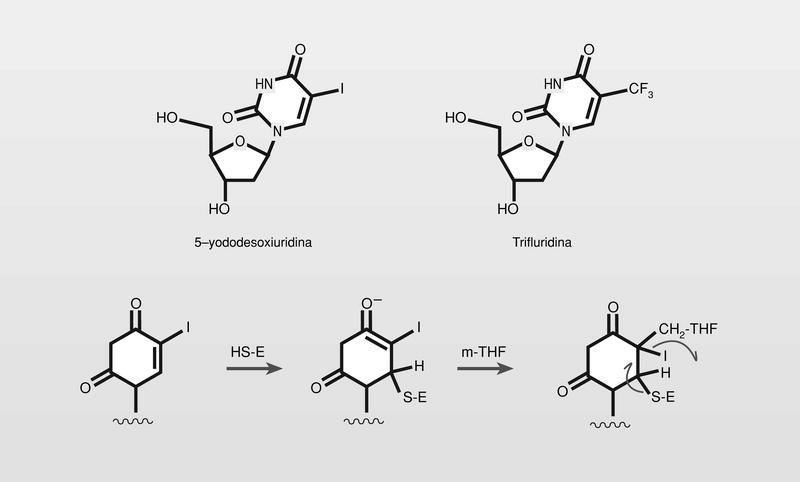
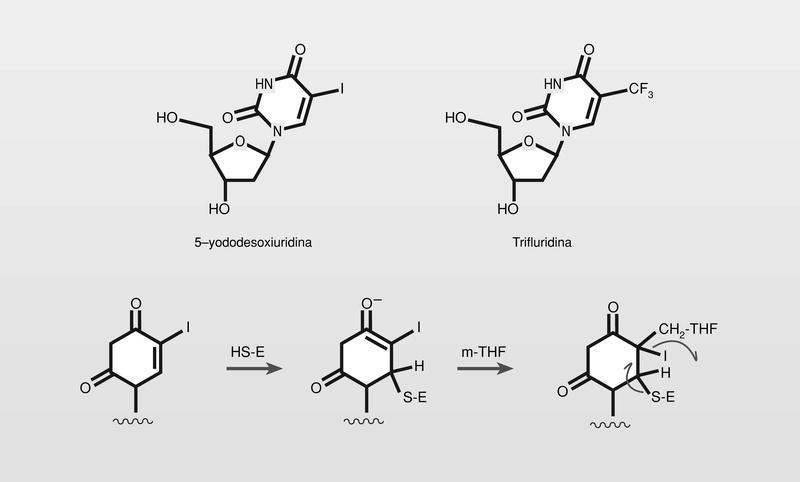
El primer análogo de nucleósido antiviral utilizado en terapia fue la 5-yododesoxiuridina (fig. 11). Es un análogo estructural de la desoxitimidina en el que el grupo metilo de esta se sustituye por un átomo de yodo (radio de Van der Waals del metilo y del yodo: 2,00 y 2,15 Å, respectivamente), y por lo tanto actúa como su antimetabolito, fosforilándose e incluyéndose en el ácido nucleico que se está formando de manera errónea. En la actualidad se utiliza en el tratamiento tópico de queratoconjuntivitis producida por herpes- virus humano (HVH) la trifluridina (5-trifluormetildesoxiuridina) (fig. 11), que sigue siendo un análogo directo de los citados (radio de Van der Waals del grupo trifluormetil, 2,44 Å). Este compuesto es más activo que la 5-yododesoxiuridina puesto que al ser el grupo trifluormetil peor grupo saliente que el yodo, bloquea en mayor medida la timidino sintetasa (fig. 11), pero mantiene el principal problema de la falta de selectividad (especificidad), puesto que tampoco su activación es estrictamente dependiente de la quinasa del virus, y afecta entonces en gran medida a la célula huésped.
Fue importante el descubrimiento de que el 5-etil derivado, es decir, un homólogo superior de la desoxitimidina, presentaba cierta actividad citotóxica en células infectadas por el HVH y sin embargo, no en células sanas. Fue una confirmación de que las quinasas virales eran menos específicas (más fáciles de engañar, es decir, fosforilan más fácilmente sustratos diferentes de aquellos normales) que las quinasas de las células huésped. Esto permite extraer una conclusión importante: para conseguir un antimetabolito antiviral lo más eficaz (con el mayor índice terapéutico) posible, es necesario apartarse lo más posible de la estructura del metabolito.
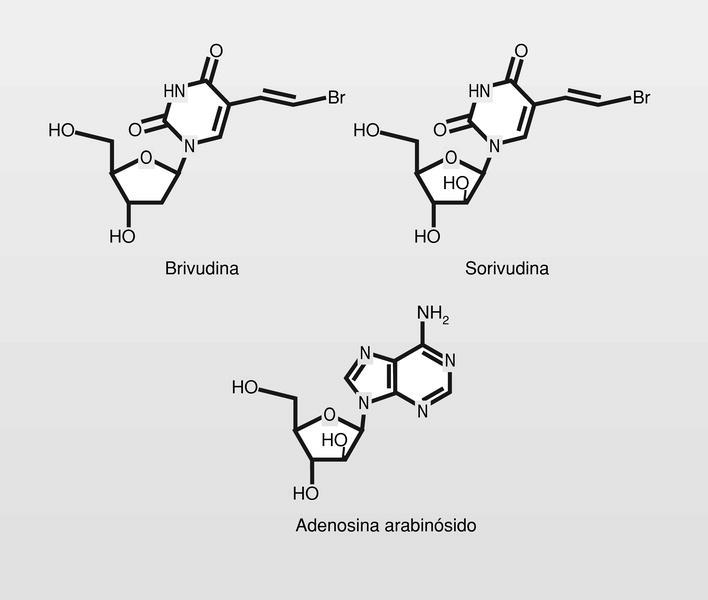
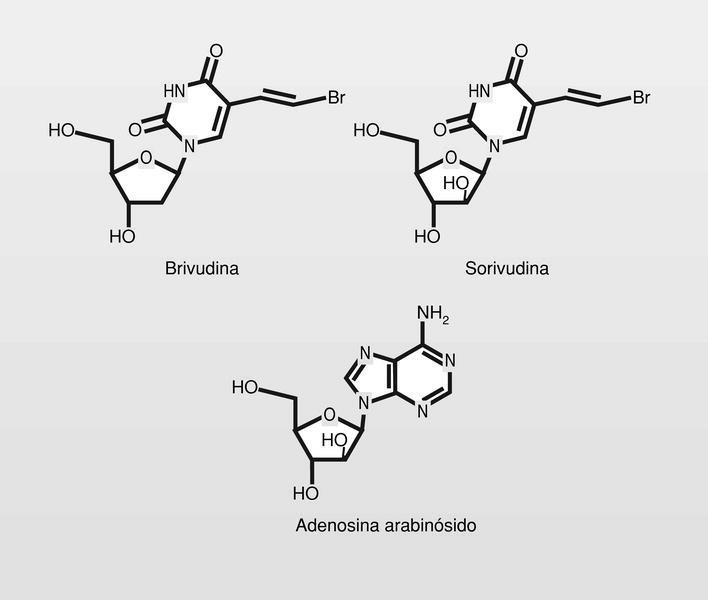
Se prepararon así, la brivudina, con un grupo bromovinilo en la posición 5 del uracilo, y más recientemente la sorivudina (análogo arabinósido) (fig. 12), que está en fase clínica avanzada para el tratamiento de HVH. Este es el más potente inhibidor conocido de varicela-zoster-virus (VZV). En los primeros ensayos clínicos del fármaco sucedieron varios casos de muerte aparentemente relacionadas con el uso del mismo. Posterior investigación detectó que los pacientes muertos habían sido tratados simultáneamente con 5-fluorouracilo (5-FU, antitumoral). El problema está en que el 5-FU se metaboliza mediante la uracil reductasa y la sorivudiva se hidroliza primero liberando el 5-bromoviniluracilo, que es un potente inhibidor de la uracilo reductasa, por lo que los niveles del altamente citotóxico 5-FU resultaban exageradamente aumentados (es preciso llamar la atención sobre estos aspectos en las situaciones de combinación de fármacos).
La adenosina arabinósido (fig. 12) fue el primer fármaco antiviral utilizado para uso sistémico. Tiene baja biodisponibilidad oral por lo que se administraba por vía intravenosa y se degrada (hidrólisis del grupo amino de la posición 6 del anillo de purina) muy rápidamente por la adenosina deaminasa.
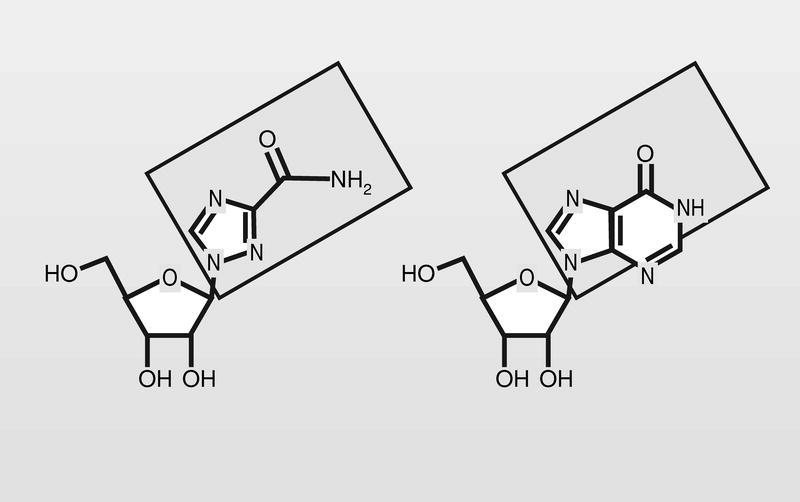
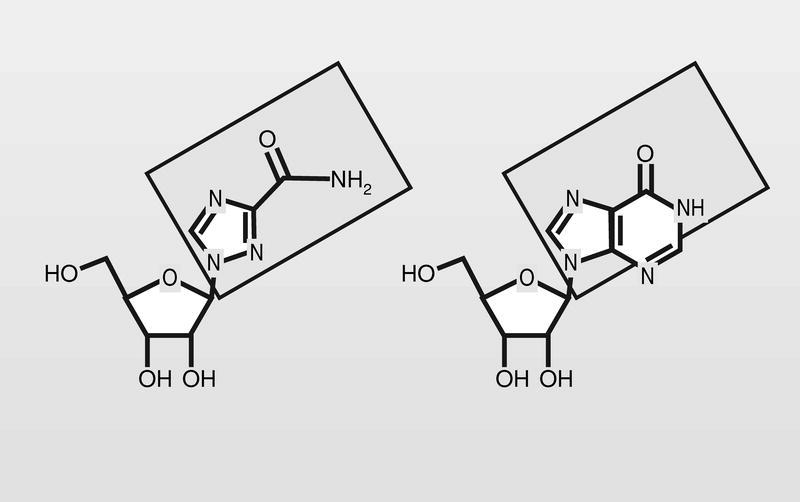
Entre los análogos de nucleósido que presentan una modificación drástica del esqueleto de la base heterocíclica, tiene un lugar destacado la ribavirina (fig. 13), con un anillo de triazol sustituido por un grupo carboxamido en la posición 3 del mismo, que es la apropiada para mimetizar las posiciones 5 y 6 de la inosina (hipoxantina). Su mecanismo de acción parece más complejo que el de otros análogos y aún no perfectamente conocido.

Cuando este sustrato se fosforila, su monofosfato inhibe la inosina monofosfato (IM) deshidrogenasa, que es necesaria para la conversión de IM en xantina monofosfato, la cual se requiere para la síntesis de guanosina trifosfato (GTP). Por otra parte, su trifosfato puede inhibir ARN polimerasas virales. Es un compuesto que muestra amplia actividad frente a gran variedad de virus ARN y ADN, pero con un específico uso en el tratamiento del virus sincitial respiratorio (VSR).
El descubrimiento del aciclovir (1974, aunque su uso clínico se desarrolló en los años 80) fue de un gran impacto porque resultó el primer inhibidor selectivo de HVH, de tal manera que disminuye drásticamente su actividad frente a otros tipos de viriasis, pero también es esencialmente inerte para las células del huésped.
La diana molecular es el ADN polimerasa viral, que confunde el trifosfato de aciclovir con dGTP y lo incorpora a la cadena de ADN viral que está formando. Como en su estructura falta el grupo hidroxilo de la posición 3', bloquea la posterior prolongación de la cadena de ADN, considerándose un «terminador de cadena» (esto establece una adicional seguridad: este análogo nunca puede formar parte, como una mutación, de un ADN maduro).
La activación del aciclovir necesita tres fosforilaciones sucesivas, que realizan tres quinasas. La pri mera debe de convertir el aciclovir en su monofosfato y es una quinasa viral, que está ausente en las células no infectadas.
El gran perfil de seguridad que ofrece este fármaco lo lleva a ser muy utilizado sobre todo en pacientes inmunodeprimidos, trasplantados, personas afectadas de cáncer, otras infecciones en personas con sida, presentando incluso un interés profiláctico.
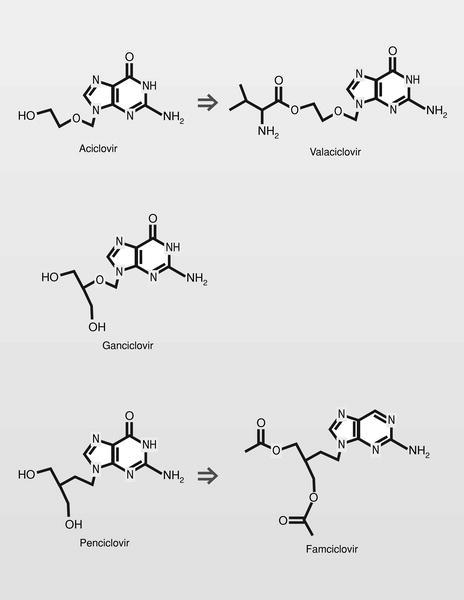
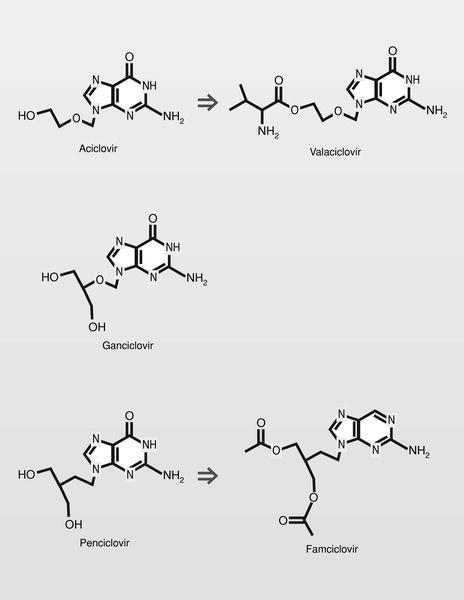
El descubrimiento y el desarrollo del aciclovir estimuló mucho el esfuerzo en el área de los aciclonucleósidos, obteniéndose varios éxitos posteriores (fig. 14):
Ganciclovir. Con un perfil farmacológico bastante distinto del aciclovir a pesar de su similitud estructural, ya que es activo frente a citomegalovirus (CMV) y mucho menos selectivo (con mucho menor margen terapéutico), produciendo fundamentalmente problemas de supresión de médula ósea.
Penciclovir. Su perfil farmacológico vuelve a ser más parecido al aciclovir, a pesar de ser un paso más en la diferenciación estructural. El trifosfato de penciclovir tiene una larga vida media, por lo que su administración puede ser menos frecuente que en el caso del aciclovir.
Existen también los profármacos, con un interés fundamental de biodisponibilidad:
Valaciclovir (L-valil éster del aciclovir), tiene la ventaja de que cuando se administra por vía oral, se hidroliza muy rápidamente en aciclovir y el aminoácido esencial L-valina, y los valores del fármaco en plasma alcanzan 3-5 veces el nivel de la administración del fármaco solo.
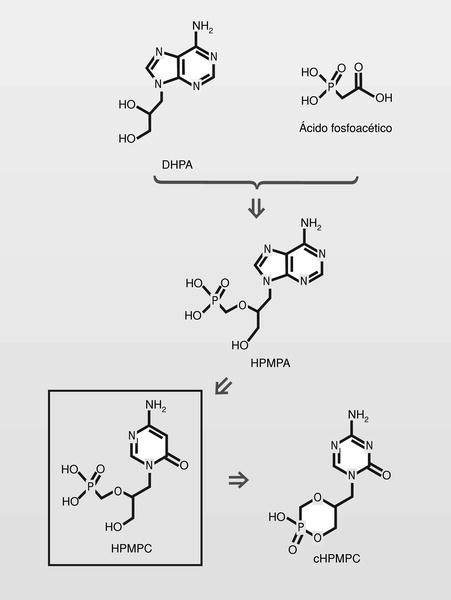
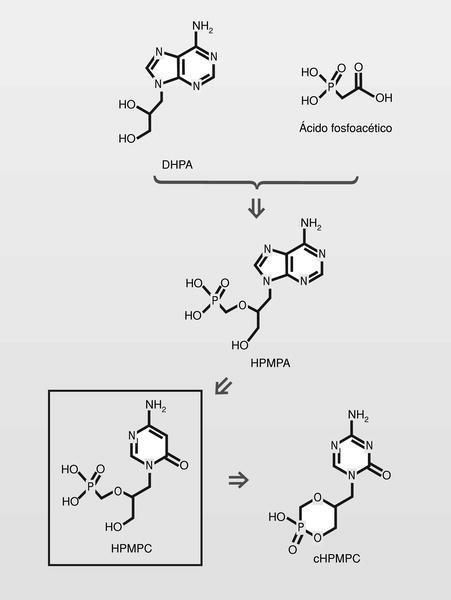
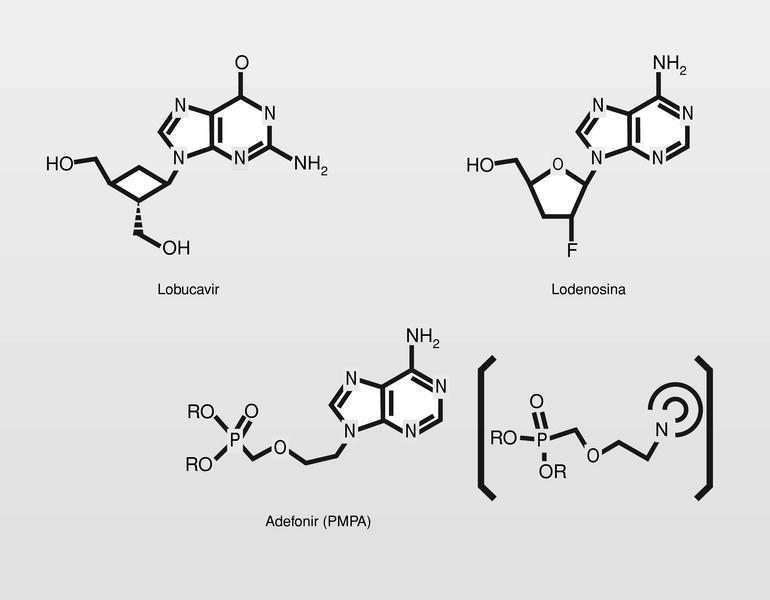
Famciclovir (diacetil éster del penciclovir). En 1978 se publica un nuevo nucleósido, la (S)-9-(2,3-dihidroxipropil) adenina (DHPA), que es el primer aciclonucleósido con amplio espectro antiviral. Muestra ser inhibidor de la (S) adenosilhomocistein-hidrolasa, y por lo tanto particularmente inhibidor de virus DNA, que necesitan hacer metilaciones dependientes de la S-adenosilmetionina. Por conjugación estructural entre este compuesto y los fosfonatos (en concreto el ácido fosfonoacético, homólogo superior del foscarnet) se llega a una nueva clase de antivirales, los fosfonatos de aciclonucleósidos (fig. 15).
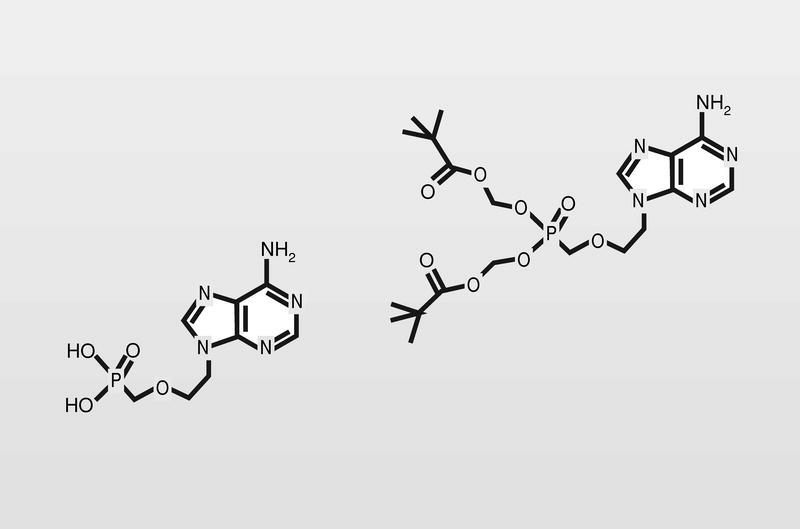
El HPMPC (cidofovir) puede usarse frente a CMV, HVH y virus papilloma humano (VPH), y tiene una duración de acción muy larga por su alta vida media. El cidofovir no sería un estricto terminador de cadena, pero sí lo es en la práctica puesto que la introducción de dos consecutivos establece tal tensión de cadena, que impide su prolongación. Están en fase de estudio clínico análogos, que sí serianterminadores de cadena estrictos (fig. 16).
Nucleósidos inhibidores de la transcriptasa inversa (NRTI)
En 1983 se conoció que el sida era causado por un retrovirus. El primer éxito en su tratamiento fue el uso del análogo de nucleósido AZT (zidovudina, 3'-azido-3'-diseoxitimidina), que había sido sintetizado 20 años antes como potencial antitumoral.
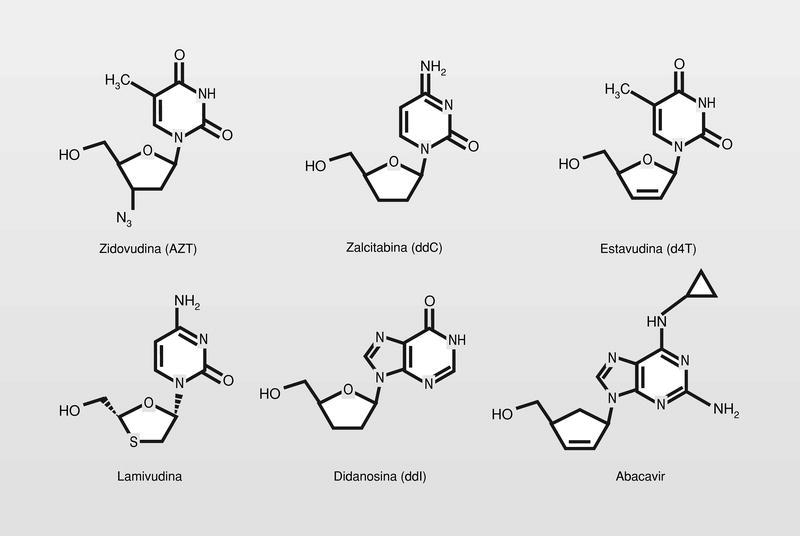
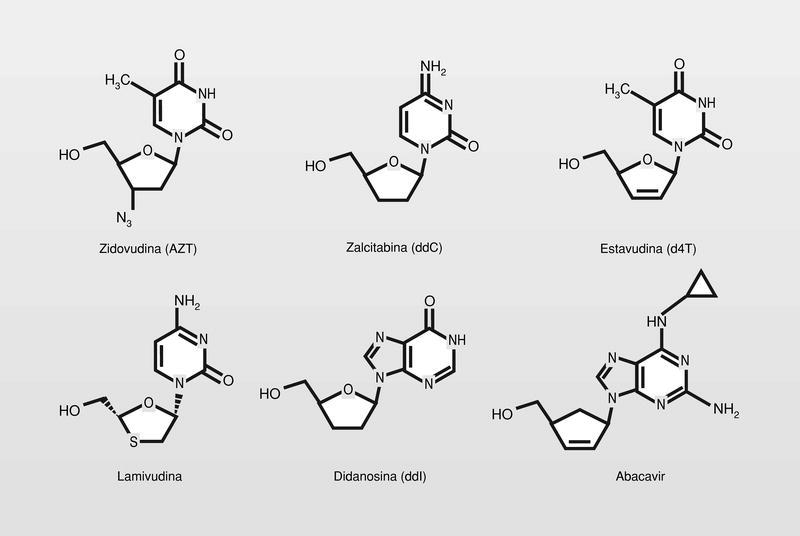
Todos los utilizados en clínica actualmente son 2',3'-dideoxinucleósidos (fig. 17) y actúan de una manera muy similar. Se mono-, di- y trifosforilan intracelularmente, de esta manera engañan a la polimerasa y al ser incluidos en el ácido nucleico que se está formando, actúan como terminadores de la cadena al no poder prolongarse la misma por carecer del grupo hidroxilo de la posición 3'.
El AZT entonces, como el aciclovir, es también un terminador de cadena, pero la mayor diferencia entre ambos modos de acción es en el ámbito de la activación por las quinasas. Mientras que el aciclovir es dependiente de una enzima codificada por el virus para su primera fosforilación, el AZT es activado en todos sus pasos por enzimas de la célula huésped. Este proceso de activación convierte al AZT en un fármaco mucho menos selectivo y con menor margen de seguridad. El AZT manifiesta solo una especificidad modesta por la transcriptasa inversa, aunque en este punto es conveniente hacer notar que en cualquier caso, el efecto es más drástico sobre el genoma viral que sobre el genoma de la célula huésped, porque esta última tiene un sistema de control celular más desarrollado que lleva a detectar y reparar con más eficacia el daño, de cuanto pueden hacerlo los organismos más simples.
Esta mayor falta de especificidad se traduce en la aparición de efectos indeseables cuando se prolonga el tratamiento (anemia y neutropenia fundamentalmente), pero el mayor problema fue, de todas maneras, la aparición de resistencias. Esto puede reducirse con fármacos alternativos que puedan ser usados en combinación, y la experiencia en el campo de antimicrobianos sugería que la mejor asociación sería aquella en la que los componentes atacasen diferentes dianas farmacológicas.
Sin embargo, como los primeros fármacos disponibles para el sida después del AZT fueron otros 2',3'dideoxinucleósido análogos y su diana la TI, combinaciones entre estos fueron las primeras ensayadas. De estas combinaciones, tres cosas me parecen especialmente destacables y sorprendentes:
Cuando el virus se hacía resistente al AZT, mantenía su sensibilidad frente a ddC y ddI (por lo tanto, no se producía resistencia cruzada incluso en fármacos que tienen como diana la misma enzima).
Cuando se utiliza la combinación se produce un efecto sinérgico sobre las actividades individuales.
El VIH en un paciente tratado con AZT adquiere mutaciones en el gen de la TI que le incrementa la resistencia al AZT. Si se cesa el tratamiento con AZT y se continúa con un nucleósido alternativo, resulta la posterior adquisición de una posterior mutación que le confiere resistencia a este segundo nucleósido, pero suprime las «lesiones inducidas por AZT» y se produce reversión de la sensibilidad a AZT (fenómeno de supresión de resistencia).
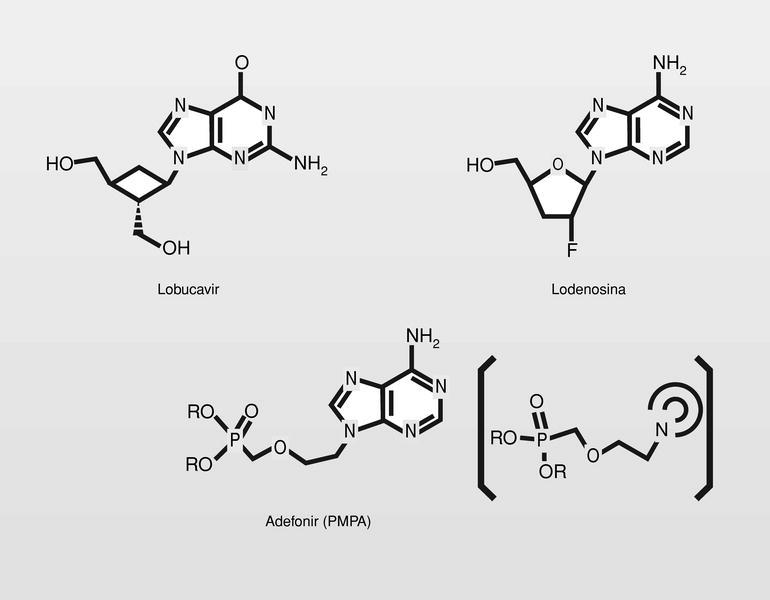
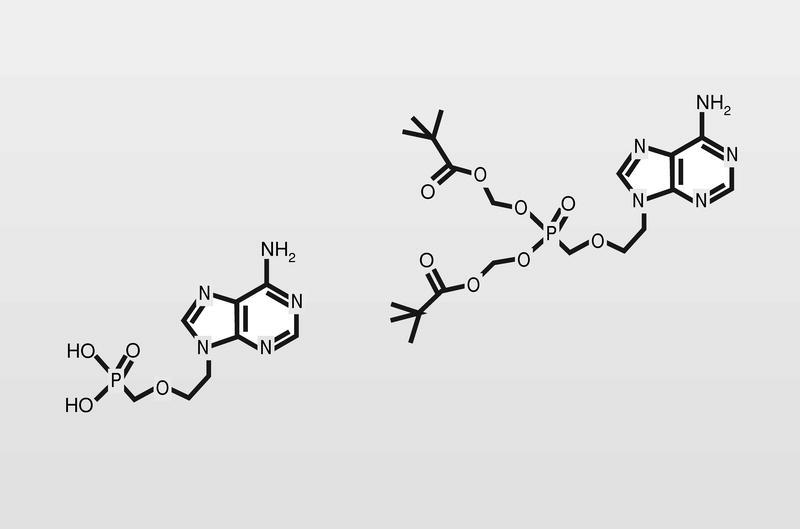
Por otro lado, se intenta mejorar la selectividad de estos análogos estudiando moléculas lo más alejadas posible del modelo para tratar de no engañar a la ADN polimerasa celular. Así están en fase clínica de estudio derivados de ciclobutano, derivados con sustituciones por átomos de flúor o fosfonatos de isósteros de aciclonucleósidos (fig. 18).
No nucleósidos inhibidores de la transcriptasa inversa (NNRTI)
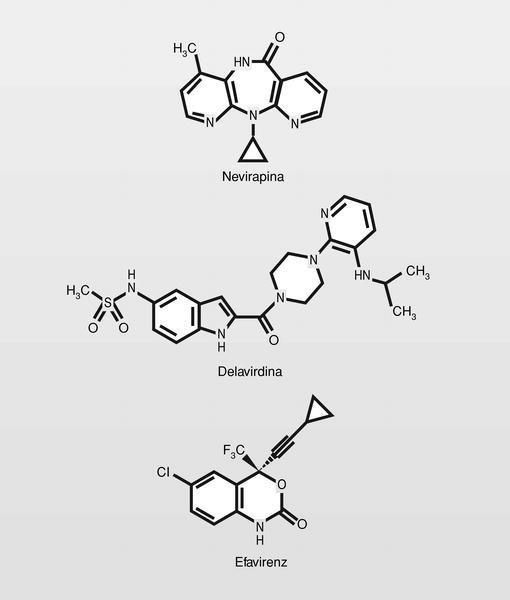
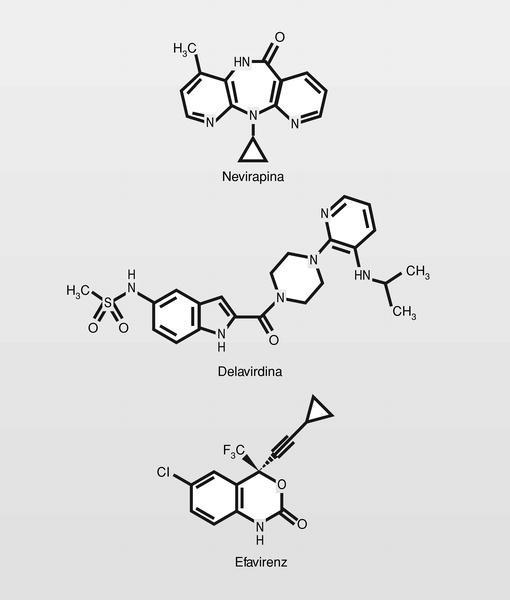
Los primeros compuestos inhibidores de la TI de este tipo fueron los denominados HEPT y TIBO a principios de los años noventa. Estos compuestos interfieren con la primera de las funciones de la TI [a) transcripción del ARN viral a ADN para formar híbridos ARN.ADN; b) degradación de la cadena de ARN, y c) replicación del ADN monocatenario] uniéndose de manera «no competitiva» en un sitio diferente (subunidad p66) del sitio catalítico, pero espacial y funcionalmente relacionado con el sitio de unión del nucleótido trifosfato. De esta manera el nucleótido trifosfato podría unirse a la enzima, pero el resultado es que no puede incluirlo en la cadena de ácido nucleico, posiblemente porque el NNRTI fuerza una estereoquímica «no eficaz» en la enzima.
Se utilizan en clínica 3 compuestos (fig. 19). Todos ellos presentan estructuras muy diferentes pero que pueden caracterizarse por una disposición de dos estructuras lipófilas en «alas de mariposa» con un cuerpo central hidrófilo7.
Terapia antisentido
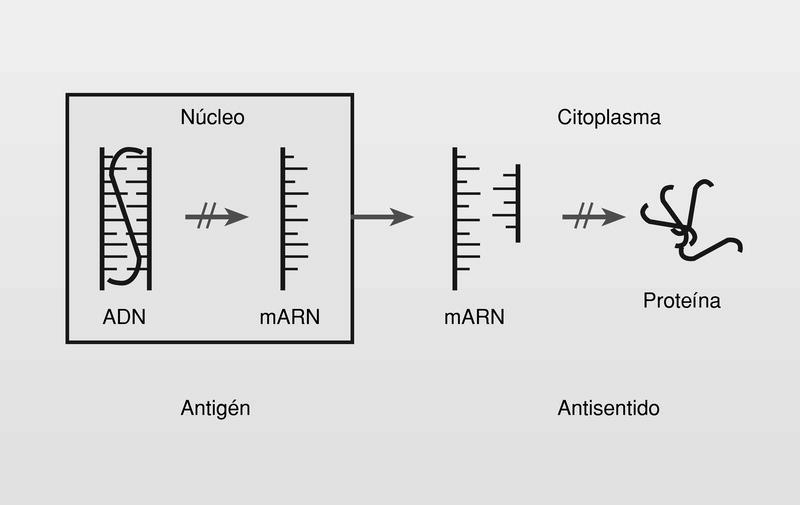
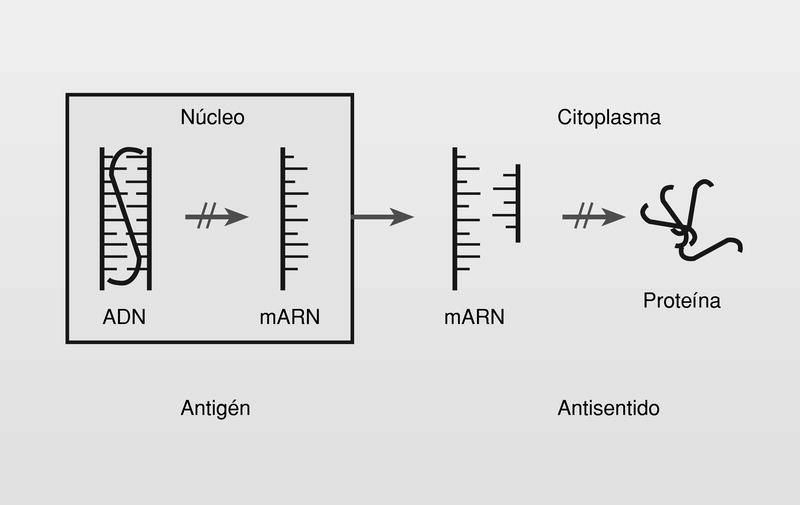
Se conoce como terapias antigén y antisentido a aquellas estrategias terapéuticas que nos permiten realizar un bloqueo de la función del ADN en el núcleo o del ARNm en el citoplasma de las células respectivamente, por acoplamiento de una nueva hebra de oligonucleótido o análogo (fig. 20). La terapia antisentido es más factible porque el oligo no tiene que penetrar en el núcleo celular3a.
El acoplamiento de la nueva hebra o cadena se basa en los mismos principios por los cuales la naturaleza regula la información genética: la complementariedad de los pares de bases A-T o A-U y G-C.
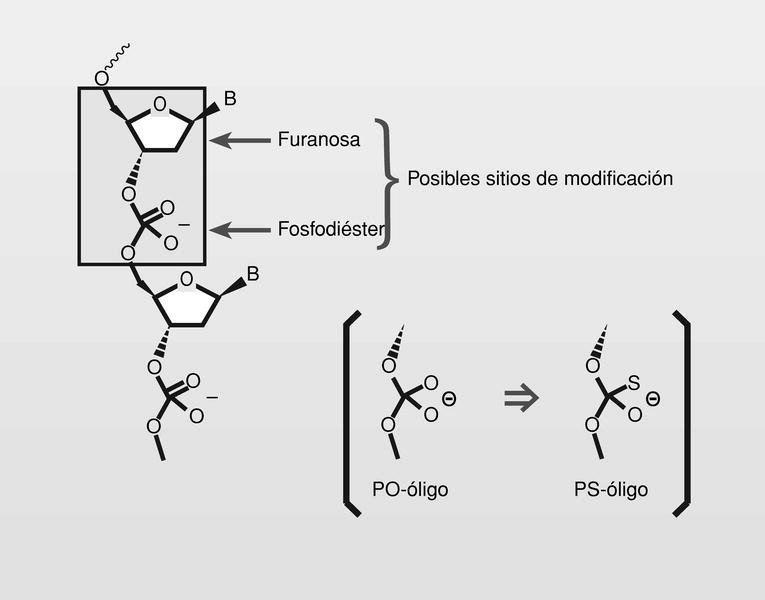
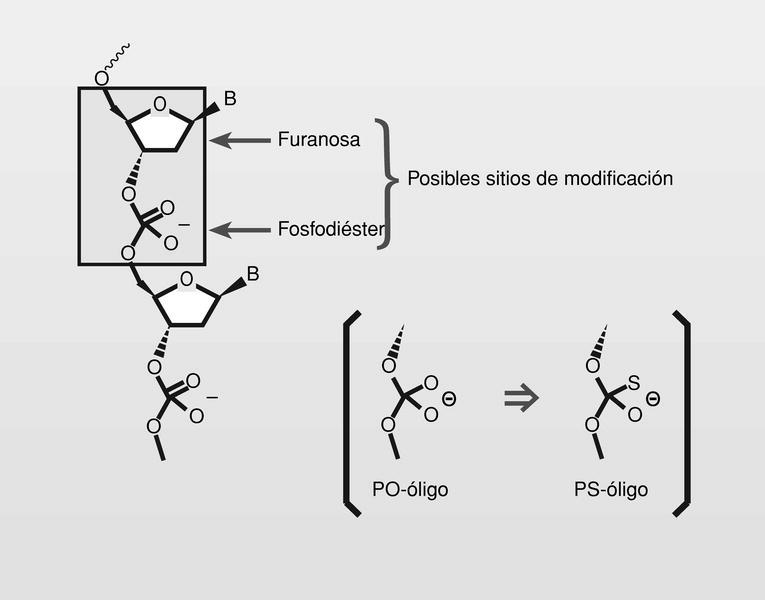
Los oligonucleótidos no modificados tienen importantes problemas para ser usados en terapéutica: poca permeabilidad celular, fácil degradación por nucleasas e incluso baja afinidad por secuencias de RNA. Esto lleva a pensar en la posibilidad de modificación de oligonucleótidos naturales bien sea en las porciones correspondientes a los anillos de furanosa o bien sea en los enlaces fosfodiéster5c. La modificación con fosforotioato (PS-oligos) resulta de la sustitución de (P = O) por (P = S) (fig. 21).
Estos son los que pueden considerarse de primera generación y a este grupo pertenece el único fármaco actualmente en uso clínico como antiviral, el fomivirsen (fig. 22), que funciona como hebra complementaria del mARN de CMV en la región 2, que codifica proteínas de regulación.
El átomo de azufre es solo ligeramente más grande que el átomo de oxígeno y al ser menos electronegativo, deja una menor carga positiva sobre el átomo de fósforo, siendo así más dificilmente degradable por las nucleasas. Los PS-oligos mostraron un efecto antisentido global superior a ninguna otra de las modificaciones realizadas.
A pesar de estas ventajas de los PS-oligos sobre los PO-oligos, ni la estabilidad frente a nucleasas, ni la biodisponibilidad fueron óptimas y, sin embargo, son propensos a unirse de forma no selectiva a diferentes proteínas, lo que les confiere los efectos colaterales. Para superar los problemas y no perder la potencia ya alcanzada con los PS-oligos, se está trabajando en los llamados «oligo mixtos» (MBO, mixed backbone oligos)8, que contienen más de un tipo de modificación de esqueleto, aunque una parte se mantiene de PS-oligo. Podemos considerarlos oligos de segunda generación y la elección de estas modificaciones y su posición en el MBO es clave para tratar de conseguir las propiedades antisentido deseadas. Se consiguen de esta manera oligos más seguros y también oligos en los que la absorción está facilitada porque se reduce la carga global de estas moléculas, con porciones de análogos casi neutros como los poliamida-ácidos nucleicos (PNA), o con guanidinas o tioureas sustituyendo al grupo fosfodiéster, lo que le confiere cargas positivas a pH ~7 por lo que junto a las negativas otorgadas por los PS-oligos, son moléculas con carácter zwiteriónico y casi carga global neutra.
Interferones
Los interferones (adoptan su nombre de «interferencia») son citocinas (cyto-célula y kinesis-movimiento): proteínas solubles en agua o glicoproteínas que libera una población celular como mediador intercelular o molécula de señalización, es decir, moléculas que son capaces de producir y liberar determinadas células para interrelacionarse con otras.
Los interferones son citocinas de bajo peso molecular (aproximadamente 20.000, alrededor de 145 aa.) producidas por muchas células eucariotas como respuesta a muchos agentes inductores.
Se descubren en 19579 y los más conocidos e importantes son: IFN-*, que se producen por infeccion viral de leucocitos; IFN-ß, que se producen por infección viral de fibroblastos; IFN-*, que se produ ce por intervención de un antígeno sobre células T (también se llama interferón «inmunitario»); y los interferones * y *, que son producidos por la placenta.
Pueden tener entonces diferentes actividades farmacológicas entre las que destacan como más importantes la antiviral y la antitumoral. Los más interesantes como antivirales son los IFN-* (que son en realidad 20 familias, siendo los utilizados en el tratamiento de la hepatitis C el IFN-*2a y el IFN-*2b) y los IFN-*. Estos son además los más parecidos entre sí, aunque el ß es una glicoproteína y, sin embargo, el * es una proteína.
Los interferones se producen en una célula cuando es infectada por un virus y se liberan protegiendo a otras células de la infección (las células sanas casi no tienen interferón puesto que este ha de ser inducido), o mejor, impiden que el virus pueda realizar su ciclo reproductivo en ellas (el interferón interfiere con la multiplicidad viral). Aunque este mecanismo de acción está suficientemente contrastado, no está descartado que pudiese tener adicionalmente algún tipo de influencia en la liberación del virus de las células infectadas o incluso en los procesos de entrada, por interacción del interferón con receptores de membrana de las células.
Las características fundamentales que los hacen interesantes como fármacos son las siguientes:
Son muy potentes y de amplio espectro (a diferencia de un anticuerpo que produce una inmunidad muy específica), es decir, no son específicos para el virus, pero sí lo son para la especie que los produce.
Son muy seguros, exentos prácticamente de toxicidad por su alta selectividad en la acción antiviral.
Los mayores problemas que presentan para ser utilizados como fármacos son:
Como todas las proteínas, son inestables (aunque el IFN-* y -* resisten bien el pH ácido y temperaturas de hasta 40-50 ºC) y tienen baja biodisponibilidad por problemas de absorción, de unión a otras proteínas plasmáticas y de fácil biodegradación
No tienen actividad frente a células infectadas, o tienen muchísima menos, por lo que, en este sentido serían más interesantes como preventivos que como curativos.
Es difícil y caro obtenerlos puros, aunque ahora es más sencillo al utilizar técnicas de ADN recombinante.
Precisamente, para tratar de minimizar estos problemas se realiza un amplio esfuerzo en el desarrollo de fármacos que pudiesen actuar como «inductores de interferón», trabajando en dos frentes:
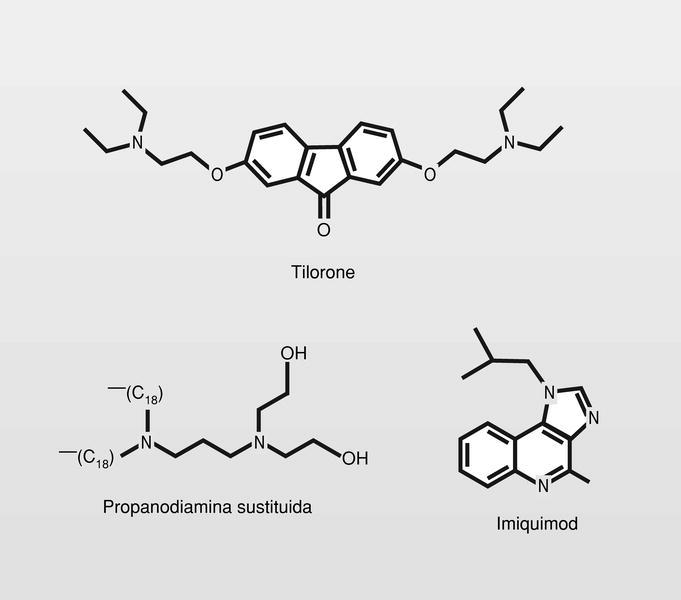
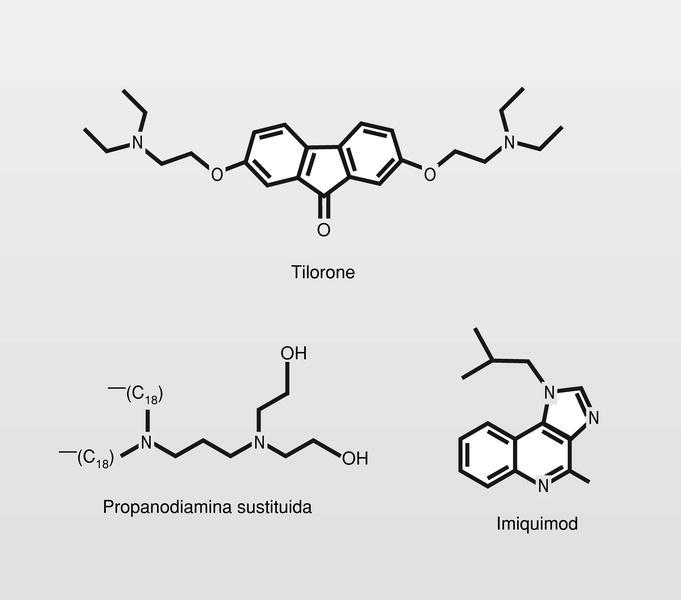
Pequeñas moléculas. Fue experimentado el tilorone que es un buen inductor en ratón, pero no así en humanos. Están en fase de experimentación clínica diversas propanodiaminas substituidas. Fue ya introducido en clínica recientemente el imiquimod, para el tratamiento de verrugas genitales externas producidas por VPH (fig. 23). Este fármaco forma parte entonces, de un nuevo grupo que puede llamarse «modificadores de la respuesta inmunitaria», desprovisto de efectos antivirales directos.
Polímeros. Se trata de polirribonucleótidos de doble hélice. Hasta el momento merece especial mención el poli I:C, que es un polirribonucleótido formado por una cadena con la base de inosina y la complementaria, con base de citosina. Para hacerlo más resistente a la hidrólisis, se estabiliza complejándolo con poli L-lisina y carboximetilcelulosa (poli ICLC). *
Bibliografía y notas
1. Considero de utilidad para aspectos generales de virus: a) Ingraham JL, Ingraham CA. Introducción a la microbiología. Barcelona: Reverté, 1998, y b) Prescott LM, Harley JP, Klein DA. Microbiología. México: McGraw-Hill Interamericana, 1999.
2. On drugs and therapeutics. The Medical Letter 2000; 5-11. Para infecciones virales no causadas por HIV (páginas 17-22 para infección por HIV). Fármacos concretos se encuentran en Drugs of the future.
3. Considero de utilidad para aspectos generales de terapia antiviral: a) Bastow KF, Akanitapichat P. Antiviral agents. En: J. Wiley. Burger's Medicinal Chemistry and Drug Discovery. Vol. 4. 5.a ed., 1997, y b) Blair E, Darby G, Gough G, Littler E, Rowlands D, Tisdale M. Antiviral therapy. Berlín: Bios Scientific Publishers. Springer-Verlag, 1998.
4. En 1952, Renato Dulbecco (Premio Nobel 1975) utilizó por primera vez un cultivo celular para el crecimiento de virus.
5. De Clercq E. Antiviral therapy for human immunodeficiency virus infections. Clin Microbiol Rev 1995; 8 (2): 200-239 (a: página 204 y referencias citadas; b: páginas 218-220, y c: páginas 217-218).
6. Kim ChU, Lew W, Williams MA, Liu H, Zhang L, Swaminathan S, Bischofberger N, Chen MS, Mendel DB, Tai ChY, Laver WG, Stevens RC. Influenza neuraminidase inhibitors possessing a novel hidrophobic interaction in the enzyme active site: design, synthesis, and structural analysis of carbocyclic sialic acid analogues with potent anti-influenza activity. J Am Chem Soc 1997; 119: 681-690. Una recopilación interesante de este artículo con bibliografía adicional, la realiza Schnaar RL. Chemtracts-Biochemistry and Molecular Biology 1998; 11: 222-228.
7. De Clercq E. Non-nucleoside reverse transcriptase inhibitors (NNRTIs) for the treatment of human immunodeficiency virus type 1 (HIV-1) infections: strategies to overcome drug resistance development. Med Research Rev 1996; 16 (2): 125-157.
8. Zhou W. Mixed-backbone oligonucleotides as second generation antisense drugs. Drugs of the Future 2000; 25 (2): 183-191.
9. Isaacs A, Lindenmann J. Prod R Soc 1957; B147: 258-263.