



Infertility is a clinical disorder affecting approximately 15% of reproductive-aged couples worldwide. Recently, the influence of oxidative stress (OS) in decreased semen quality has been discussed. OS corresponds to an imbalance between oxidants and antioxidants defenses, present in the organism. High levels of reactive oxygen species (ROS) damage biomolecules present in sperm cells and may lead to the loss of membrane integrity, DNA fragmentation or even to death by apoptosis. This study aimed to evaluate the correlation between human semen clinic parameters and parameters that assessed the presence of OS.
Material and methodsA total of 32 semen samples, obtained from a randomized group of donors, were included in this study. Basic semen parameters were analyzed according to the WHO's guidelines. The total antioxidant capacity of sperm cells was measured as well as the expression of certain antioxidant proteins, namely superoxide dismutase (SOD) and glutathione peroxidase 4 (GPx4), by colorimetric techniques and immunoblotting, respectively. The effect of ROS in spermatozoa protein oxidation was analyzed by determining the presence of 3-nitrotyrosine and carbonyl groups, by slot blot. Lipid peroxidation was evaluated, by performing the thiobarbituric acid reactive substances (TBARS) assay with colorimetric tests.
ResultsThe results indicated that SOD was negatively correlated with viscosity (p=0.035), volume (p=0.004) and carbonyl groups presence (p=0.005). This protein also showed a positive correlation with the presence of tail defects in sperm cells (p=0.044). In turn, GPx4 showed a negative correlation with the presence of non-progressive motile spermatozoa (p=0.012). TBARS assay revealed a negative correlation with the concentration of sperm cells (p=0.000) and the total number of spermatozoa (p=0.026), but, in turn, this assay showed a positive correlation with the volume of semen (p=0.038).
ConclusionIt was concluded that the seminal quality is affected by the oxidative balance in sperm cells. The obtained results suggest that lipid peroxidation leads to a reduction in sperm concentration; antioxidant proteins protect the spermatozoa against protein oxidation and contribute to an increased sperm motility and normal semen viscosity. Thus, evaluation of oxidative parameters may be a useful tool for male infertility diagnosis and follow-up of antioxidant treatments.
A infertilidade é um problema clínico que afeta cerca de 15% dos casais em idade fértil a nível mundial. Nos últimos anos, a influência do stress oxidativo (SO) em amostras de sémen com qualidade reduzida tem sido discutida. O SO corresponde a um desequilíbrio entre as espécies oxidantes e as defesas antioxidantes presentes no organismo. Níveis elevados de espécies reativas de oxigénio (ROS) danificam as biomoléculas presentes nos espermatozoides e podem provocar perda da integridade da membrana, fragmentação de DNA ou até apoptose. Este estudo teve como objetivo avaliar a correlação entre os parâmetros clínicos do sémen humano e os parâmetros avaliadores da presença de SO.
Material e métodosUm total de 32 amostras de sémen, obtidas a partir de um grupo aleatório de dadores, foi incluído neste estudo. Os parâmetros seminais básicos foram analisados de acordo com os padrões da OMS. A capacidade antioxidante total dos espermatozoides e a expressão das proteínas antioxidantes SOD e GPx4 foram avaliadas por técnicas colorimétricas e immunoblotting, respetivamente. O efeito das ROS na oxidação proteica dos espermatozoides foi avaliado pela determinação da presença de grupos carbonilo e de 3-nitrotirosina, por slotblot. A peroxidação lipídica foi avaliada pela técnica dos TBARS, por testes colorimétricos.
ResultadosOs resultados indicaram que a SOD estava negativamente correlacionada com a viscosidade (p=0,035), o volume (p=0,004) e a presença de grupos carbonilo (p=0,005), e positivamente correlacionada com a presença de anomalias na cauda dos espermatozoides (p=0,044). A GPx4 demonstrou uma correlação negativa com a presença de espermatozoides móveis não progressivos (p=0,012). A técnica dos TBARS revelou uma correlação negativa com a concentração (p=0,000) e o número de espermatozoides (p=0,026) e uma correlação positiva com o volume de sémen (p=0,038).
ConclusãoConcluiu-se que a qualidade seminal é afetada pelo equilíbrio oxidativo nos espermatozoides. Os resultados obtidos sugerem que a peroxidação lipídica leva à redução da concentração de espermatozoides; os antioxidantes protegem os espermatozoides contra a oxidação proteica, melhorando também a mobilidade e alterando a viscosidade do sémen. Assim, sugerimos a avaliação dos parâmetros oxidativos como uma nova ferramenta útil para o diagnóstico e monitorização dos tratamentos da infertilidade masculina.
Infertility is classified by the World Health Organization (WHO) as a “disease of the reproductive system defined by the failure to achieve a clinical pregnancy after 12 months or more of regular unprotected sexual intercourse”.1 Infertility is a common clinical situation that affects around 15% of the reproductive-aged couples worldwide, of which around 50% of the cases are due to male factors.2,3 In turn, around half of the male infertility cases still have idiopathic causes. However, the increase of oxidative stress (OS) has been considered a possible etiology for this pathological situation. OS is defined as an imbalance between the levels of reactive oxygen species (ROS) and the levels of antioxidant systems, where ROS are in excess.3–7
Sources of reactive oxygen speciesLevels of ROS may be increased due to its production by endogenous and exogenous sources. In fertile men, the ejaculate is usually constituted by several cell types, such as mature spermatozoa, immature sperm cells, leukocytes and epithelial cells.5,7,8 Among these, leukocytes (particularly neutrophils and macrophages) and abnormal or immature spermatozoa are the principal endogenous sources of ROS.7,9,10 Similarly, several environmental and lifestyle factors also modulate the levels of ROS in seminal fluid. For example, tobacco smoke has more than four thousand chemical compounds, of which many are ROS and reactive nitrogen species (RNS). These ROS and RNS of tobacco smoke cause higher rates of DNA sperm fragmentation and axoneme damages as well as a decreased concentration of spermatozoa in semen.3,4,11
Reactive oxygen species functionAt optimal concentrations, ROS play an important physiologic role, regulating fertilization, acrosome reaction, hyperactivation, motility and capacitation.4,5,12,13 However, when the levels of ROS are increased, they damage spermatozoa's biomolecules, including lipids, proteins and DNA.
The spermatozoon plasma membrane is rich in polyunsaturated fatty acids (PUFA), which makes it vulnerable to oxidation by ROS.4,14,15 Phospholipids peroxidation promotes alterations in membrane fluidity, leading to an alteration in sperm motility parameters. A byproduct of lipid peroxidation is malondialdehyde (MDA), which is commonly analyzed in laboratories to measure peroxidative damage in spermatozoa.4,16,17 ROS can also damage mitochondrial and nuclear DNA in human spermatozoa, which occurs due to the action of ROS on phosphodiester backbones and DNA bases, leading to its fragmentation.18,19
Another effect of ROS is the decrease in the number of sperm cells, through activation of the apoptotic process. Alterations in the mitochondrial membrane fluidity lead to apoptosis, due to the release of cytochrome c and the apoptosis inducing factor (AIF) into the cytosol, with consequent activation of caspases.20,21
ROS are also responsible for protein oxidation, inducing enzymatic activity or structural protein function impairment. Although the oxidized proteins are functionally inactive and quickly removed, some may accumulate in the organism and be the cause of several pathologies.5,11,22
Elevated levels of nitric oxide can induce protein nitration in tyrosine residues. This process corresponds to the addition of a NO2 group at the third position of either free or protein-bound tyrosine, leading to the formation of 3-nitrotyrosine (3-NT).23 Nitric oxide and superoxide anion (O2•−) react with each other, in the presence of CO2, producing peroxynitrite (ONOO−), which is highly reactive. The reaction of this molecule with proteins can result in S-nitrosylation or in the production of 3-NT.23–25
Antioxidant defensesThe human organism has antioxidant defense systems, which, in the ejaculate, are localized in seminal plasma and spermatozoa.26 These include, for instance, superoxide dismutase (SOD) that catalyzes O2•− conversion into H2O2 and O2 and has different isoforms. SOD2, conjugated with manganese in its active site (MnSOD), is localized in the mitochondrial matrix and acts in O2•− produced in mitochondria.27 SOD1 is conjugated with copper and zinc (CuZnSOD), and localizes in the cytoplasm.28 There is another isoform of SOD, extracellular SOD (also called SOD3), which is produced by Sertoli cells and germ cells. Glutathione peroxidase (GPx) is a selenoenzyme that catalyzes the reduction of H2O2 to H2O, with the concomitant oxidation of glutathione.11 Glutathione peroxidase is localized in mitochondria, nucleus and acrosome region of spermatozoa.26 This protein possesses several isoforms; however, the most abundant isoform in sperm cells is GPx4.29
Diagnosis of male infertilityThe diagnosis of male infertility is usually made by basic semen analysis, which evaluates several parameters of seminal quality such as concentration, morphology and motility of sperm cells as well as volume, appearance, viscosity, pH and liquefaction of semen.30 Further, to complement these results, several tests can be performed to evaluate the etiology of the infertility, but it is not a common practice in clinical setting.31,32
In this study, we evaluated the total antioxidant capacity, the expression of specific antioxidants (SOD and GPx4) and protein oxidation and lipid peroxidation caused by ROS effect (namely, 3-nitrotyrosine, TBARS and carbonyl groups formation) in the proteins and lipids of the sperm cells, in order to address how these parameters correlate with basic semen analysis. These assays may help to understand the etiology of idiopathic male infertility and the alterations observed in the basic semen analysis. Our aim is to suggest the application of new assays in the diagnosis and in the follow-up of fertility treatments.
MethodsSample collection and basic semen analysisA total of 32 semen samples were obtained from a randomized group of donors (fertile and infertile men, healthy and with infertility-related diseases such as varicocele) that attended the Urology service of the Hospital Infante D. Pedro, by masturbation to a sterile container, after 3–5 days of sexual abstinence. Semen samples were analyzed according to the WHO's laboratory manual guideline.30 After complete liquefaction of the semen samples at 37°C, during approximately 30min, the macroscopic examination was performed by evaluating the liquefaction, viscosity, appearance and volume of the semen samples. The microscopic examination included the analysis of motility, concentration and morphology of the spermatozoa. Further, sperm motility was analyzed and its classification was based on three groups: motile progressive, motile non-progressive and immotile spermatozoa, as recommended by the WHO. The concentration of sperm cells was determined by counting them, after proper dilution, using an improved Neubauer chamber. The results were expressed as number of sperm cells per milliliter. The sperm morphology was then assessed by preparing a smear of semen, followed by fixation with methanol and staining using a hemacolor kit (Merck, Germany). The morphology was calculated based on the percentage of normal vs abnormal forms, and the abnormalities can refer to head, middle piece and tail defects as well as an excess of residual cytoplasm.
Sample preparationAfter semen liquefaction, spermatozoa were separated from seminal plasma by centrifugation (600×g for 5min at 4°C) and supernatant (seminal plasma) was frozen at −20°C and stored. The pellet obtained, which corresponds to the spermatozoa, was washed and then centrifuged two more times with PBS. Some of the aliquots of the resulting pellet were directly frozen at −20°C, while the others aliquots were resuspended in 1% SDS, before being frozen.
Protein assayProtein concentration was measured using bicinchoninic acid (BCA) assay (Pierce Biotechnology, USA). This method is based on the reduction of Cu2+ to Cu+, which is promoted by proteins in an alkaline medium (the biuret reaction); colorimetric detection of Cu+ cation is performed using a unique reagent containing BCA. The chelation of two molecules of BCA with Cu+ ion is responsible for the formation of a purple-colored reaction product, which exhibits a strong absorbance at 562nm that is linear with increasing protein concentrations over a working range of 20–2000μg/mL. In this assay, protein standards containing 0, 1, 2, 5, 10 and 20μg of bovine serum albumin (BSA) were used to obtain a standard curve. Three microliters of each sample with 22μL of 1% SDS were added to each well of a microplate, followed by the addition of 200μL of working reagent. The microplate was incubated for 30min at 37°C and, after cooling to room temperature, the absorbance was measured at 562nm in a microplate reader (TECAN, Genius, Männedorf, Switzerland).
Total antioxidant statusThe concentration of antioxidant proteins in sperm cells was measured using a test for total antioxidant status (TAS) determination (Randox, Crumlin, UK). This test is based on the incubation of ABTS with a peroxidase and H2O2, which produces a radical cation ABTS+ with a stable blue-green color, measured at 600nm. The addition of the sperm cells samples may cause a suppression of the color formation due to the presence of antioxidants. The protocol performed was optimized in our laboratory. The modified protocol was based on the total protein concentration, namely 15μg, and the cuvette method was modified to a microplate method. In this protocol, 5μL of samples were mixed with 200μL of chromogen (metmyoglobin/ABTS reagent) and the reaction was initiated by adding 40μL of H2O2. The final concentration was measured at 600nm after incubation for 3min at 37°C.
Detection of antioxidant enzymes by Western blottingThe protein expression levels of the antioxidant enzymes SOD and GPx4 were evaluated by running sperm samples onto 15% SDS-PAGE gels followed by Western blot using appropriate antibodies. Briefly, 50μg of protein extract for each sample was prepared in 40μL of total volume and applied into the wells and allowed to run at 90mA. A protein marker (NZYColour Protein Marker II) was also applied for molecular weight comparison and to check the efficiency of the transfer.
Proteins separated by SDS-PAGE were electrotransferred to a nitrocellulose membrane. The non-specific binding sites of the membrane were blocked in 1× TBS-T/5% low fat milk for 1h. The membrane was then incubated for 2h with the primary antibodies anti-β-tubulin, anti-SOD1 and anti-GPx4 (Merck Millipore, Germany). All antibodies were diluted 1:1000 in 1× TBS-T/5% low fat milk. After that, the membrane was washed three times with 1× TBS-T during 10min. Then, the membrane was incubated for 1h with the respective fluorescent secondary antibodies, anti-Rabbit and anti-Mouse (LI-COR BioSciences), diluted 1:5000 in 1× TBS-T/5% low fat milk: anti-mouse for β-tubulin and SOD1 and anti-rabbit for GPx4. Finally, the membrane was washed as before and scanned in a LI-COR's Odyssey Infrared Imaging System (LI-COR Biosciences).
Carbonyl groups (CG) determinationTo determine the carbonyl groups, the slot blot method followed by immunodetection were performed. Initially, the samples was prepared with an amount of 30μg of protein per 50μL of sample. Fifty microliters of 12% SDS was then added, followed by the addition of 100μL of 20mM DNPH/10% trifluoroacetic acid (TFA). After an incubation of 30min in the dark, the solution was neutralized with 75μL of 2M Tris/18% of mercaptoethanol. Then, the samples were diluted with 1× PBS to a concentration of 2ng/μL and 100μL of each sample were applied in the slot-blot and transferred to a nitrocellulose membrane, which was then incubated in 10% methanol for activation and washed in water and 1× TBS-T. The membrane was blocked for 1h in 1× TBS-T/5% low fat milk and then incubated with the rabbit antibody anti-DNP (Merck KGaA, Darmstadt, Germany) and anti-mouse secondary antibody. Then, the membrane was washed with 1× TBS-T and scanned using a Li-cor's Odissey Infrared Imaging System.
3-Nitrotyrosine determinationTo determine the presence of 3-nitrotyrosine (3-NT), the samples of spermatozoa were diluted in 1× TBS, to obtain a final concentration of 1ng/μL. A volume of 100μL was then pipetted into a nitrocellulose membrane, using the slot-blot technique. This membrane was then permeabilized with 10% methanol and washed with water and 1× TBS-T. The membrane was blocked with 5% milk in 1× T-TBS and incubated with primary antibody anti-nitrotyrosine (Merck KGaA, Darmstadt, Germany) (1:1000) and secondary antibody anti-rabbit (1:5000). At last, the membrane was washed two times with 1× T-TBS and revealed in a Li-cor's Odissey Infrared Imaging System.
Thiobarbituric acid reactive substance (TBARS) assayThe method for the determination of TBARS was based on Ohkawa protocol.33 In this assay, 200μL of sample was added to 400μL of trichloroacetic acid (TCA) 10%, followed by a centrifugation during 20min at 1500×g. Four hundred microliters of 1% thiobarbituric acid (TBA) were then added to 400μL of supernatant. After that, the samples were placed in a boiling water bath during 10min. The samples were then cooled, in cold water, for approximately 20min. Finally, the absorbance was measured at 535nm in a spectrophotometer.
Statistical analysisThe statistical analysis was conducted using the SPSS 22.0 software.34 First, a descriptive statistics to each quantitative parameter analyzed was performed. To evaluate the relation between the results obtained during the seminal basic analysis and the oxidative balance analysis (namely total antioxidant capacity, antioxidant expression and protein oxidation) a Spearman's rank correlation analysis was performed. The Spearman's rank correlation coefficient is a nonparametric measure of statistical dependence between two variables.
The semen samples were divided into two groups (normal and abnormal samples according to the standards of the WHO) and then the Shapiro–Wilk test and Levene's test for equality of variance were performed. After validation of the statistical assumptions and assuming that the samples are independent and homoscedastic, T-Student test for equality of means was performed. A significance level of 5% was used.
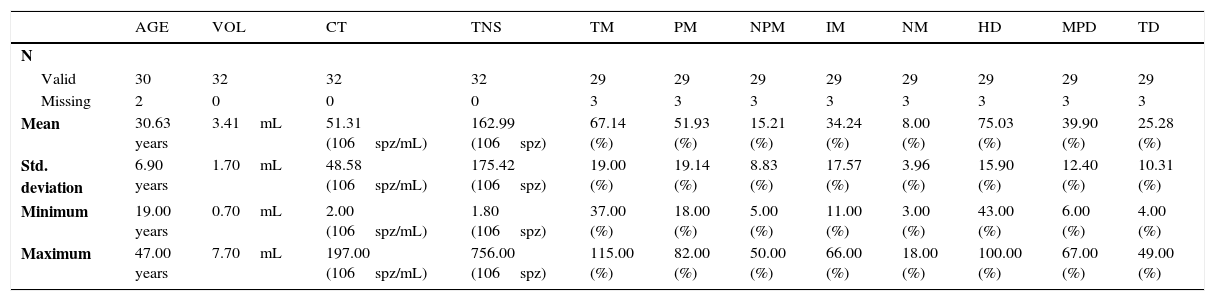
ResultsHuman semen samples (n=32) were subjected to a basic semen analysis and, then, descriptive statistics of the results obtained was performed (Table 1).
Descriptive statistics of basic semen analysis.
| AGE | VOL | CT | TNS | TM | PM | NPM | IM | NM | HD | MPD | TD | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | ||||||||||||
| Valid | 30 | 32 | 32 | 32 | 29 | 29 | 29 | 29 | 29 | 29 | 29 | 29 |
| Missing | 2 | 0 | 0 | 0 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Mean | 30.63 years | 3.41mL | 51.31 (106spz/mL) | 162.99 (106spz) | 67.14 (%) | 51.93 (%) | 15.21 (%) | 34.24 (%) | 8.00 (%) | 75.03 (%) | 39.90 (%) | 25.28 (%) |
| Std. deviation | 6.90 years | 1.70mL | 48.58 (106spz/mL) | 175.42 (106spz) | 19.00 (%) | 19.14 (%) | 8.83 (%) | 17.57 (%) | 3.96 (%) | 15.90 (%) | 12.40 (%) | 10.31 (%) |
| Minimum | 19.00 years | 0.70mL | 2.00 (106spz/mL) | 1.80 (106spz) | 37.00 (%) | 18.00 (%) | 5.00 (%) | 11.00 (%) | 3.00 (%) | 43.00 (%) | 6.00 (%) | 4.00 (%) |
| Maximum | 47.00 years | 7.70mL | 197.00 (106spz/mL) | 756.00 (106spz) | 115.00 (%) | 82.00 (%) | 50.00 (%) | 66.00 (%) | 18.00 (%) | 100.00 (%) | 67.00 (%) | 49.00 (%) |
CT: concentration; VOL: volume; TNS: total number of spermatozoa; TM: total motility; PM: progressive motile spermatozoa; NPM: non-progressive motile spermatozoa; IM: immotile; NM: normal morphology; HD: head defects; MPD: middle piece defects; TD: tail defects; SPZ: spermatozoa.
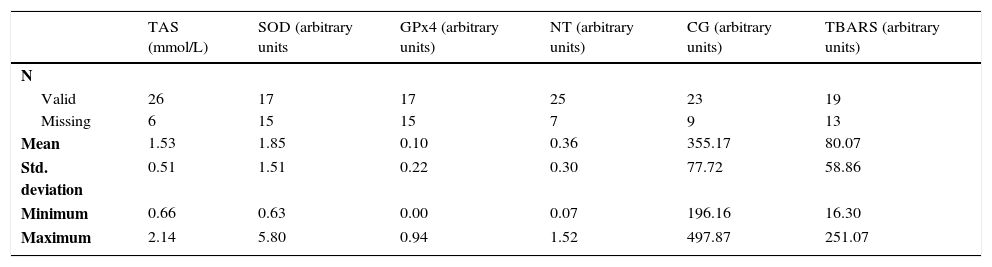
Sperm extracts were prepared and assayed in order to determine the total antioxidant status (TAS). Also, the expression of the antioxidant proteins SOD and GPx4 was quantified by Western blotting. Further, protein oxidation (carbonyl groups and 3-NT) and lipid peroxidation (TBARS) were determined by slot-blot technique. The results of these assays were initially analyzed by performing descriptive statistics (Table 2).
Descriptive statistics of the assays performed.
| TAS (mmol/L) | SOD (arbitrary units | GPx4 (arbitrary units) | NT (arbitrary units) | CG (arbitrary units) | TBARS (arbitrary units) | |
|---|---|---|---|---|---|---|
| N | ||||||
| Valid | 26 | 17 | 17 | 25 | 23 | 19 |
| Missing | 6 | 15 | 15 | 7 | 9 | 13 |
| Mean | 1.53 | 1.85 | 0.10 | 0.36 | 355.17 | 80.07 |
| Std. deviation | 0.51 | 1.51 | 0.22 | 0.30 | 77.72 | 58.86 |
| Minimum | 0.66 | 0.63 | 0.00 | 0.07 | 196.16 | 16.30 |
| Maximum | 2.14 | 5.80 | 0.94 | 1.52 | 497.87 | 251.07 |
CG: carbonyl groups; Gpx4: glutathione peroxidase 4; 3-NT: 3-nitrotyrosine; SOD: superoxide dismutase; TAS: total antioxidant status; TBARS: thiobarbituric acid reactive substances.
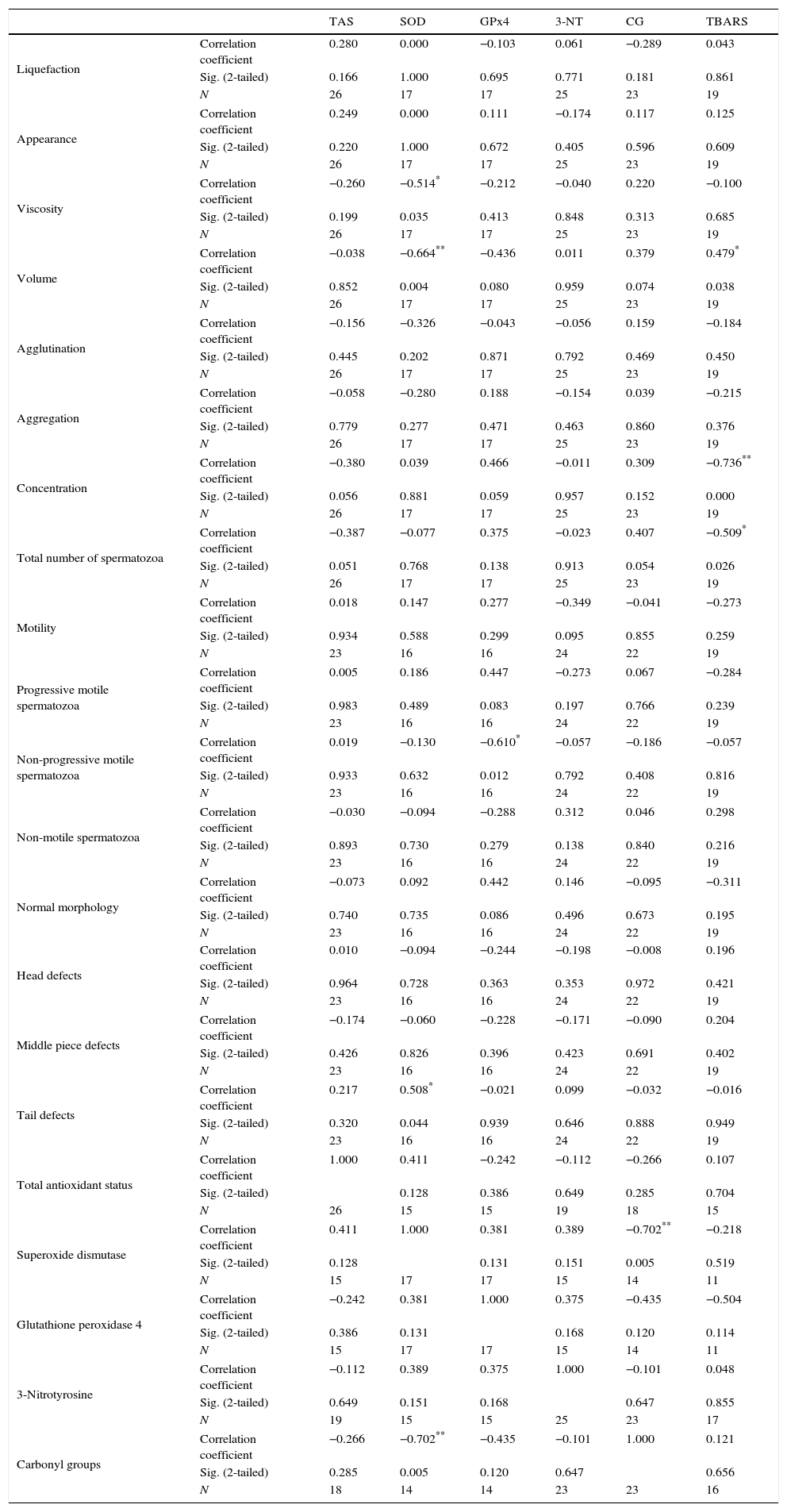
After a Normality test and a visual inspection of the box-plots it was decided to use a nonparametric correlations method. Thus, the correlation between the parameters of the basic semen analysis and the assays performed was evaluated by performing a Spearman's rho correlation, rs (Table 3).
Spearman's rho correlation between seminal parameters and the oxidative balance of sperm cells.
| TAS | SOD | GPx4 | 3-NT | CG | TBARS | ||
|---|---|---|---|---|---|---|---|
| Liquefaction | Correlation coefficient | 0.280 | 0.000 | −0.103 | 0.061 | −0.289 | 0.043 |
| Sig. (2-tailed) | 0.166 | 1.000 | 0.695 | 0.771 | 0.181 | 0.861 | |
| N | 26 | 17 | 17 | 25 | 23 | 19 | |
| Appearance | Correlation coefficient | 0.249 | 0.000 | 0.111 | −0.174 | 0.117 | 0.125 |
| Sig. (2-tailed) | 0.220 | 1.000 | 0.672 | 0.405 | 0.596 | 0.609 | |
| N | 26 | 17 | 17 | 25 | 23 | 19 | |
| Viscosity | Correlation coefficient | −0.260 | −0.514* | −0.212 | −0.040 | 0.220 | −0.100 |
| Sig. (2-tailed) | 0.199 | 0.035 | 0.413 | 0.848 | 0.313 | 0.685 | |
| N | 26 | 17 | 17 | 25 | 23 | 19 | |
| Volume | Correlation coefficient | −0.038 | −0.664** | −0.436 | 0.011 | 0.379 | 0.479* |
| Sig. (2-tailed) | 0.852 | 0.004 | 0.080 | 0.959 | 0.074 | 0.038 | |
| N | 26 | 17 | 17 | 25 | 23 | 19 | |
| Agglutination | Correlation coefficient | −0.156 | −0.326 | −0.043 | −0.056 | 0.159 | −0.184 |
| Sig. (2-tailed) | 0.445 | 0.202 | 0.871 | 0.792 | 0.469 | 0.450 | |
| N | 26 | 17 | 17 | 25 | 23 | 19 | |
| Aggregation | Correlation coefficient | −0.058 | −0.280 | 0.188 | −0.154 | 0.039 | −0.215 |
| Sig. (2-tailed) | 0.779 | 0.277 | 0.471 | 0.463 | 0.860 | 0.376 | |
| N | 26 | 17 | 17 | 25 | 23 | 19 | |
| Concentration | Correlation coefficient | −0.380 | 0.039 | 0.466 | −0.011 | 0.309 | −0.736** |
| Sig. (2-tailed) | 0.056 | 0.881 | 0.059 | 0.957 | 0.152 | 0.000 | |
| N | 26 | 17 | 17 | 25 | 23 | 19 | |
| Total number of spermatozoa | Correlation coefficient | −0.387 | −0.077 | 0.375 | −0.023 | 0.407 | −0.509* |
| Sig. (2-tailed) | 0.051 | 0.768 | 0.138 | 0.913 | 0.054 | 0.026 | |
| N | 26 | 17 | 17 | 25 | 23 | 19 | |
| Motility | Correlation coefficient | 0.018 | 0.147 | 0.277 | −0.349 | −0.041 | −0.273 |
| Sig. (2-tailed) | 0.934 | 0.588 | 0.299 | 0.095 | 0.855 | 0.259 | |
| N | 23 | 16 | 16 | 24 | 22 | 19 | |
| Progressive motile spermatozoa | Correlation coefficient | 0.005 | 0.186 | 0.447 | −0.273 | 0.067 | −0.284 |
| Sig. (2-tailed) | 0.983 | 0.489 | 0.083 | 0.197 | 0.766 | 0.239 | |
| N | 23 | 16 | 16 | 24 | 22 | 19 | |
| Non-progressive motile spermatozoa | Correlation coefficient | 0.019 | −0.130 | −0.610* | −0.057 | −0.186 | −0.057 |
| Sig. (2-tailed) | 0.933 | 0.632 | 0.012 | 0.792 | 0.408 | 0.816 | |
| N | 23 | 16 | 16 | 24 | 22 | 19 | |
| Non-motile spermatozoa | Correlation coefficient | −0.030 | −0.094 | −0.288 | 0.312 | 0.046 | 0.298 |
| Sig. (2-tailed) | 0.893 | 0.730 | 0.279 | 0.138 | 0.840 | 0.216 | |
| N | 23 | 16 | 16 | 24 | 22 | 19 | |
| Normal morphology | Correlation coefficient | −0.073 | 0.092 | 0.442 | 0.146 | −0.095 | −0.311 |
| Sig. (2-tailed) | 0.740 | 0.735 | 0.086 | 0.496 | 0.673 | 0.195 | |
| N | 23 | 16 | 16 | 24 | 22 | 19 | |
| Head defects | Correlation coefficient | 0.010 | −0.094 | −0.244 | −0.198 | −0.008 | 0.196 |
| Sig. (2-tailed) | 0.964 | 0.728 | 0.363 | 0.353 | 0.972 | 0.421 | |
| N | 23 | 16 | 16 | 24 | 22 | 19 | |
| Middle piece defects | Correlation coefficient | −0.174 | −0.060 | −0.228 | −0.171 | −0.090 | 0.204 |
| Sig. (2-tailed) | 0.426 | 0.826 | 0.396 | 0.423 | 0.691 | 0.402 | |
| N | 23 | 16 | 16 | 24 | 22 | 19 | |
| Tail defects | Correlation coefficient | 0.217 | 0.508* | −0.021 | 0.099 | −0.032 | −0.016 |
| Sig. (2-tailed) | 0.320 | 0.044 | 0.939 | 0.646 | 0.888 | 0.949 | |
| N | 23 | 16 | 16 | 24 | 22 | 19 | |
| Total antioxidant status | Correlation coefficient | 1.000 | 0.411 | −0.242 | −0.112 | −0.266 | 0.107 |
| Sig. (2-tailed) | 0.128 | 0.386 | 0.649 | 0.285 | 0.704 | ||
| N | 26 | 15 | 15 | 19 | 18 | 15 | |
| Superoxide dismutase | Correlation coefficient | 0.411 | 1.000 | 0.381 | 0.389 | −0.702** | −0.218 |
| Sig. (2-tailed) | 0.128 | 0.131 | 0.151 | 0.005 | 0.519 | ||
| N | 15 | 17 | 17 | 15 | 14 | 11 | |
| Glutathione peroxidase 4 | Correlation coefficient | −0.242 | 0.381 | 1.000 | 0.375 | −0.435 | −0.504 |
| Sig. (2-tailed) | 0.386 | 0.131 | 0.168 | 0.120 | 0.114 | ||
| N | 15 | 17 | 17 | 15 | 14 | 11 | |
| 3-Nitrotyrosine | Correlation coefficient | −0.112 | 0.389 | 0.375 | 1.000 | −0.101 | 0.048 |
| Sig. (2-tailed) | 0.649 | 0.151 | 0.168 | 0.647 | 0.855 | ||
| N | 19 | 15 | 15 | 25 | 23 | 17 | |
| Carbonyl groups | Correlation coefficient | −0.266 | −0.702** | −0.435 | −0.101 | 1.000 | 0.121 |
| Sig. (2-tailed) | 0.285 | 0.005 | 0.120 | 0.647 | 0.656 | ||
| N | 18 | 14 | 14 | 23 | 23 | 16 | |
CG: carbonyl groups; Gpx4: glutathione peroxidase 4; 3-NT: 3-nitrotyrosine; SOD: superoxide dismutase; TAS: total antioxidant status; TBARS: thiobarbituric acid reactive substances.
The results demonstrated some relations between seminal parameters and the assays performed. It is shown that SOD has a statistically significant correlation in negative direction with the viscosity (rs=−0.514; 15 d.f.; p=0.035), the volume (r=−0.664; 15 d.f; p=0.004) and the carbonyl groups (rs=−0.702; 12 d.f.; p=0.005) and the strength of correlations is moderate, moderate to strong and strong respectively. SOD also showed a statistically significant correlation with the presence of tail defects (rs=0.508; 14 d.f.; p=0.044), in this case, in positive direction and with moderate strength.
In turn, GPx4 showed a statistically significant negative and moderate (to strong) correlation with the presence of non-progressive motile spermatozoa (rs=−0.610; 14 d.f.; p=0.012).
The presence of TBARS is significantly correlated with the concentration of sperm cells (rs=−0.736; 17 d.f.; p=0.000) and the total number of spermatozoa (rs=−0.509; 17 d.f.; p=0.026) in negative direction and strong way. In turn, this assay showed a positive and moderated statistically significant correlation with the volume of semen (rs=0.479; 17 d.f.; p=0.038).
DiscussionAs previously mentioned, ROS promote biomolecules damage in sperm cells, such as lipids, proteins and DNA, which may affect the morphology and motility of sperm cells among others. To better understand these alterations, spermograms of a total of 32 samples from a heterogeneous clinical set, were performed. Then, to evaluate the OS in sperm cells, three oxidative parameters were evaluated: the total antioxidant capacity, the antioxidant protein expression and the lipid and protein damage in sperm cells. Our major aim was to relate the clinical analysis of sperm cells with their oxidative balance. In this study, we did not perform the complete analysis to all the samples because they do not have the necessary volume and/or were too diluted, being impossible to use the desired volume with the needed concentration to perform the assays.
As shown in Fig. 1, in this study, it was verified that SOD is the parameter with most correlations. The results showed a negative correlation between this protein and the viscosity and volume of the semen and the carbonyl groups of spermatozoa. Furthermore, it was also verified that SOD is positively correlated with tail abnormalities of the sperm cells. In turn, a negative correlation between GPx4 and non-progressive motile spermatozoa was observed. Concerning lipid peroxidation, evaluated by TBARS assay, the results showed a negative correlation with the concentration and the total number of sperm cells, but also a positive correlation with volume.

Glutathione peroxidase 4, as previously mentioned, is an antioxidant protein that leads to the reduction of H2O2 into H2O, and it is mainly found in mitochondria of sperm cells, conferring a high protection to this organelle. In this study, a negative correlation between the levels of GPx4 and the motility of spermatozoa was found. An increase in the production of ROS and, consequently, the peroxidation of mitochondrial lipids, causes abnormalities in mitochondria that can have negative effects on motility. Glutathione peroxidase 4 in mitochondria plays a major role in suppressing the effects of ROS, contributing to a better motility of sperm cells. In spermatids, GPx4 is enzymatically active, which contributes to deletion of ROS (essentially H2O2) and thus prevents the occurrence of mitochondria alterations, such as lipid peroxidation. In turn, in mature spermatozoa, GPx4 plays a structural role, preventing morphologic alterations, such as tail defects, that can decrease motility.
Remembering that semen is also composed by fluids secreted by the male accessory glands, containing several proteins that are essential to its coagulation and liquefaction, prostate or seminal vesicles pathologies can lead to hyperviscosity. Oxidative stress is considered to be indirectly a cause of hyperviscosity, due to an increase of leukocytes, which in turn lead to the referred function alteration of the accessory gland cells. Oxidative stress is represented by an increase of ROS, but also by a decrease of the antioxidant capacity. Thus, decreased levels of SOD can represent the presence of OS that in turn leads to hyperviscosity.
With regard to SOD, carbonyl groups assay showed a negative correlation with the presence of this protein. These results suggest that SOD protects the spermatozoa against ROS, preventing changes in the side chains of proteins due to the addition of the carbonyl groups.
Besides the results already discussed, some other controversial results were found: superoxide dismutase was positively correlated with abnormalities in the tail. In this study, we analyzed CuZnSOD, an isoform present in the cytoplasm of sperm cells. However, this protein was also observed with a high expression in the testis, more specifically into the spermatogonia of the seminiferous tubules, as well as into ductus deferens and prostate cells. An increase of OS in testis is usually accompanied by an increase of OS in the remaining reproductive tract. So, an increase of OS in testis leads to a high expression of CuZnSOD by spermatogonia. However, spermatocytes and spermatids have a low expression of CuZnSOD, being more susceptible to ROS effect in late spermatogenesis and thus leading to tail defects. When mature spermatozoa with ROS damages are found in epididymis, the seminal fluid donates CuZnSOD to spermatozoa to minimize the effect of ROS in sperm cells. In turn, when OS is observed in testis, which leads to an impairment of spermatogenesis, OS in the remaining reproductive tract is also observed, and consequently, ductus deferens, seminal vesicles and prostate cells increase their antioxidants production and expression. As a result, the presence of this protein in seminal fluid increases, being then transported to sperm cells.
To measure the lipid peroxidation of sperm cells we performed a TBARS assay, which evaluated the production of MDA. TBARS concentration showed a significant negative correlation with some basic routine sperm parameters, particularly the concentration of sperm cells. As previously referred, MDA is a byproduct of lipid peroxidation, which is a process that damages the sperm cell membrane, due to its high content in PUFA. In turn, lipid peroxidation is responsible for alterations in the fluidity of cell membranes and to promote a membrane fusion as well as a decrease of membrane enzymes and ionic channels activity, which may lead to a decrease of the concentration of sperm cells.
Lipid peroxidation showed a positive correlation with the volume of semen, and this finding was consistent with the results obtained in previous studies.35 The increase in semen volume is associated with an increase of abnormal sperm and leukocytes levels, which are, as previously mentioned, the major sources of ROS. These increased levels of ROS are then responsible for an increased lipid peroxidation.
At last, a strong negative correlation between SOD and ejaculate volume was observed. This correlation was already described, and there was no information to explain this result.36 In turn, another study obtained contradictory results referring that an antioxidant intake in diet leads to an increased semen volume.37 This increased volume of semen may be due to inflammation of the accessory glands, namely of the prostate.30 Consequently, this situation is responsible for the increase of leukocytes number, leading to OS that, in turn, is also represented by a decrease in the antioxidant capacity.
ConclusionIn spermatozoa, when in increased amounts, ROS cause oxidative stress, which, in turn, leads to biomolecules alterations that modifies the physiology of these cells. In this study, it was verified that there is a correlation between the oxidative parameters, namely antioxidant levels and ROS effect in proteins and lipids, with the quality of the sperm cells. Thus, it was verified that an increase in lipid peroxidation is responsible for a reduction of the concentration of sperm cells, but a correlation between this parameter and the volume of the samples was also suggested. The levels of GPx4 were correlated with non-motile progressive motility of spermatozoa, while SOD has demonstrated some correlations with volume and viscosity of the semen, the number of tail defects and the production of CG in sperm cells.
Therefore, with these results we suggest that the evaluation of oxidative parameters (namely MDA, TAS, SOD, GPx4, CG and 3-NT) may be used as a diagnostic, prognostic or follow-up tool for male infertility; however a larger population is needed to correlate our findings with specific medical conditions affecting male infertility.
Ethical disclosuresProtection of human and animal subjectsThe authors declare that no experiments were performed on humans or animals for this study.
Confidentiality of dataThe authors declare that they have followed the protocols of their work center on the publication of patient data
Right to privacy and informed consentThe authors have obtained the written informed consent of the patients or subjects mentioned in the article. The corresponding author is in possession of this document.
Conflicts of interestThe authors have no conflicts of interest to declare.
This work was supported by the Associação Portuguesa de Urologia, through the research grant called “Criação de plataforma para avaliar a fertilidade masculina”.
