


The practice of exercise is not only a manner to maintain physical performance but also affects several systems in the organism. Several recent works have demonstrated that physical activity affects metabolic, hormonal and immunological homeostasis in the organism. On the other hand, an equilibrated diet is one of the main factors to be taken into consideration in physical performance. Metabolic syndromes severely affect muscle capacity in animal models and humans. In fact, diet and exercise, induce a similar response in organs, tissues and cells since both cause regulation of the mechanisms involved in energy regulation. Under energy challenge, organism respond regulating metabolism and reach a more balanced mechanism that affect the homeostasis of the whole body.
Maintenance of physical performance during aging is one of the main objectives to increase the quality of life in old people. By this, old people would increase their freedom decreasing their dependence, and probably their life-span but, more importantly, their health. In the present paper we compare the changes produced by both, caloric restriction (CR) and exercise in muscle performance. Furthermore, we also review the effect produced by polyphenols, bioactive compounds found in plants, that recently have been shown to importantly influence physical activity in animals. Thus, the aim of this paper is to find common mechanisms that relate energy control and muscle performance with these compounds and that can be considered as targets for future therapies against cardiovascular, metabolic diseases and age-related diseases and morbidity.
Balance of cell metabolism, a key factor for longevity and physical performanceThe concept of CR was coined at the middle of XX century as the reduction of the amount of calories ingested respecting of the average amount. From then, many different effects of CR have been described by using several organisms as models, from yeast to humans. The most interesting effect of CR is the increase in life-span found in different organisms although not yet demonstrated in humans. The increase in longevity found with CR can be due to changes in different systems including modifications of cellular bioenergetics, reduction of free radicals production from cell metabolism that can produce a delay in the appearance of diseases linked to aging.
Several works have suggested that exercise can induce similar effects than CR because the increase energy expenditure and reduction of calories available for other functions or for storage. In fact, several of the effects found in animals fed under CR have been also observed in aerobic exercise trained animals1. In experiments performed in rodents, free voluntary exercise and training were able to decrease morbidity and mortality similarly to the effect found with CR2. However, not all the studies shown to date have demonstrated a positive effect of the practice of moderate exercise in longevity3. The increase in free radical production induced by exercise would negatively affect life-span whereas the reduction in oxidative stress induced by CR would increase it3. However, several other reports demonstrate that both, CR and exercise are able to increase the cell response against oxidative stress by increasing the expression of intracellular antioxidant mechanisms. Thus, exercise and CR can finally decrease oxidative stress and increase life-span. Actually, it has been shown that moderate exercise decrease oxidative damage in old people4 in a similar way to the effect observed with CR in model animals5. Then, reduction of the amount of energy available by both, reduction of calories intake by CR or by increasing the energy expenditure by exercise seems to produce a higher resistance of the tissues against oxidative stress and a higher life expectancy.
Beside the controversial results on the effect of exercise on longevity, several other results do seem to show a closed similarity between the effect of CR and moderated and continuous aerobic exercise on cell physiology and activity in different organs and systems. Thus, the effect of CR on cardiovascular system and brain are similar to the effects found in trained organisms6. Moreover, CR and aerobic exercise synergize on the increase of muscular activity under electric stimulation in rats7. Another study have demonstrated that both, aerobic exercise and CR reduce tumour growth and sensitivity of cells against tumour inducing agents probably by affecting metabolic changes and reducing nutrient flux8. Then, it is clear that CR and exercise can positively affect several diseases and morbidity probably by inducing the same mechanisms.
Caloric restriction and aerobic exercise increase physical resistanceAlong the last years we have been working on the effect of CR and aerobic exercise on physical activity and resistance by using mice as model. Animals were fed under AL and CR (every-other-day fed model) for 6 months. During the last 2 months under these conditions a subgroup performed a training protocol consisting in running on treadmill five days a week. At the end of the experiment, animals performed different performance tests. Depending on the test, the results were different. Some of the tests demonstrated an effect of the diet and other an effect of training. We demonstrated that CR and training synergized to increase motor coordination and muscle performance to strenuous conditions. However and more surprisingly, CR, by itself, was also able to increase the resistance of animals to extenuation9.
The effect of CR on physical resistance can be due to the induction of mitochondrial biogenesis found in muscle fibers induced by CR10,11, an effect also induced by exercise12,13. Probably, both, CR and exercise, modulate the same key factors involved in mitochondrial biogenesis such as PGC1α, SIRT1, AMPK or nitric oxide (NO)14. In fact, in obese human patients, combination of CR and exercise induces mitochondrial biogenesis by activating several intracellular factors. At the same time, CR and exercise decrease whole-body oxygen consumption and DNA damage, indicating a more balanced mitochondrial activity15. Furthermore, experiments performed on cell cultures also demonstrate that serum factors from CR animals modify mitochondrial activity to a more effective activity16.
Caloric restriction and aerobic exercise decrease oxidative stress in muscleOne of the clearest effects of CR on the organism is a lower production of ROS16 and thus, a lower oxidative damage even found in muscle17. Surprisingly, both exercise and CR increase mitochondrial mass in muscle. At a first glance, this increase would increase the production of ROS18, however, mitochondria from trained animals release less amount of ROS when they are stimulated19. In fact, exercise increase the amount of oxidative type I fibers that show higher amounts of mitochondria and oxidative metabolism but show at the same time lower levels of hydrogen peroxide release and increase of antioxidant enzymes that reduce oxidative damage19.
In our experiments, trained animals fed under CR have shown lower muscle damage after being running to extenuation as determined by the levels of creatine kinase in plasma at the same time that they maintained higher levels of antioxidant activity in plasma (Rodríguez-Bíes et al, unpublished results). In the case of the other groups, the sedentary AL group suffered higher damage of muscle besides having been running less than the other groups and wasted the highest levels of antioxidant plasmatic capacity. On the other hand, when we measured levels of lipid peroxidation in muscle membranes, we found that CR decreased lipid peroxidation in muscle independently of the exercise performed by the animals. Then, CR can prevent oxidative damage induced by a strenuous exercise in muscle.
Both, CR and exercise are able to increase the expression of antioxidant enzymes in muscle20,21. This response is explained by the hormetic hypothesis that postulates that a moderated stress, induced by both, dietary stress or exercise, activates cellular defenses against oxidative stress22,23. Thus, by this mechanism, cells are more protected against a higher oxidative injury. By this mechanism, the moderate stress induced by exercise or CR will activate mitochondrial changes to induce a lower production of oxidative stress at the same time that induce the expression of antioxidant enzymes such as catalase, superoxide dismutase, glutathione peroxidases, etc. It has been already reported that this adaptive response induced against moderated oxidative stress finally produces a beneficial effect on the health of the whole organism affecting importantly muscle activity24.
The intensity of the exercise is important for the induction of antioxidant enzymes. Moderated induction of ROS during exercise is important for the induction of antioxidant enzymes by activating the activity of factors such as NF-κB and MAPK25,26. However, voluntary exercise inducing lower production of ROS seems to be insufficient to activate these factors and increase the expression of antioxidant enzymes and also induce bioenergetic changes in muscle27. Recently, it has been proposed that the free radicals generated during mild to moderate endurance-type exercise actually form part of a mechanism of adaptation to the exercise. This mechanism includes mitochondrial biogenesis, increase of muscle blood supply, and altered fuel consumption patterns increasing fat and decrease carbohydrate consumption28. It seems clear now that free radicals actually act as intracellular signalling molecules that can initiate the adaptation to exercise in a hormetic response. However, this effect of exercise not only affects muscle since, during exercise, skeletal muscle, but also liver and brain show a similar adaptive response: increased antioxidant/damage repair enzyme activity, lower oxidative damage, and increased resistance to oxidative stress indicating that exercise affects the whole organism increasing the resistance of the cells against a higher oxidative damage29. In fact, it has been recently proposed that consumption of vitamin C after exercise actually impairs the induction of exercise-dependent muscle changes due to reduction of the activation of these pathways30. However, in other studies, increase of the expression of some proteins after exercise due to supplementation with antioxidants has been also shown31. Then, the effect of antioxidants on muscle changes induced by exercise is not completely clear and can depend on the intensity and type of exercise.
Role of antioxidants in muscle performanceSeveral dietary components, including antioxidants, are currently taken into consideration as ergogenic compounds. However, to date, the role of antioxidants as ergogenic factors is not clear32 and several studies using plant extracts rich in antioxidants show equivocal results33,34.
In non-trained or in elderly people, moderate exercise improves cardiorespiratory function and reduces the levels of factors involved in cardiovascular risk. However, in these groups of people or in people living under malnutritional conditions, high oxidative injury can be induced by inadequate exercise. In these cases, antioxidant supplementation reduces oxidative damage and increases the beneficial effects of the practice of exercise35. Thus, the supplementation with antioxidants in some specific cases seems to be important to synergize with the effect of exercise.
Recently, the importance or a great group of plant compounds with bioactive capacity is increasing as antioxidant and modulators of several activities in the organism. These compounds, known as polyphenols, are uniquely characterized by the presence of more than one phenol group in the molecule. They are considered the most abundant antioxidant in the diet and are being used as the base of several reposition beverages for sport. However, the diversity of their structures makes them different from other antioxidants. This diversity permits to find compounds with very interesting therapeutic properties against cancer, cardiovascular diseases, inflammation, bioenergetic regulation and several other functions.
According to the nature of their carbon skeleton, polyphenols are classified in: phenolic acids, flavonoids, stilbenes and lignans. Flavonoids are the most abundant polyphenols in the diet because they are present ubiquitously in plants. These compounds can be classified in several different classes. According to the degree of oxidation of the oxygen heterocycle we can find: flavones (luteolin, apigenin), flavonols (quercetin), isoflavones (genistein and daidzein), anthocyanins (red fruidspigments), flavanols (catechins), proanthocyanidins (polymeric flavanols) and flavanones (hesperidin, narigin)36. Another group that is increasing its importance as antioxidant and bioactive compounds are the stilbenes. Stilbenes are also represented in nature. Among the stilbenes, the phytoalexin resveratrol have recently attracted attention for its different effects on cells, tissues, and organs. Resveratrol (3,4’,5-trihydroxystilbene), has been found in significant levels in different kind of berries, walnuts, grapes and then, red wine.
Resveratrol effect on muscle activityResveratrol is currently considered as a mimetic of CR because it affects several pathways regulated by CR and modulates bioenergetics in a similar way than CR. As CR, resveratrol supplementation of diet has increased muscle performance in mice37. This compound affects metabolic activities in the organism increasing the effectiveness of mitochondria and the antioxidant capacity of the cells. Moreover, recent results obtained in our laboratory and others have demonstrated that resveratrol increases resistance of animals to strenuous exercise37,38.
Resveratrol emerges as an important substance to protect cells against deterioration during aging. As CR, resveratrol reduces the protein expression pattern found in both cardiac and skeletal muscle during aging39. Very recently, the use of resveratrol and aerobic exercise has also demonstrated positive effects on delay of physical decline during aging40. Thus, the use of resveratrol as dietary supplement during aging can be considered to maintain an active lifestyle in old people.
Resveratrol is able to increase the glucose uptake in skeletal muscle41. The effect of resveratrol is dependent on the activation of AMPK and GLUT4 transporters41,42. Resveratrol can also increase the proliferation of muscle precursor cells (MPC) positively affecting the regeneration of muscle after injury43. MPC show impaired ability to regrow and regenerate nucleus during aging. Overexpression of the NAD(+)-dependent histone deacetylase SIRT1, one of the main targets of resveratrol, increased MPC proliferation and cell cycle progression. In fact, activation of SIRT1 with resveratrol increases MPC proliferation while inhibition of this protein with nicotinamide lowered proliferation. These findings could have medical significance since MPC proliferation is implicated in the regulation of skeletal muscle growth, its maintenance, and repair and the aging-related loss of skeletal muscle mass43.
Another important effect of resveratrol depends on its antioxidant effect on tissues. It has been demonstrated that resveratrol can prevent oxidative damage in muscle after ischemia-reperfusion44 as determined by decrease of levels of lipid peroxidation (MDA), carbonyl and protein sulphydryl levels in quadriceps muscle. Then, resveratrol can protect the skeletal muscle tissue against global ischemia and reperfusion injury because of its strong antioxidant and cytoprotective properties45. Polyphenols are regarded as xenobiotics by the organism and then, activate the expression of phase I and phase II detoxifying enzymes including enzymes involved in antioxidant protection. In fact, some polyphenols have been shown to increase the expression of gamma-glutamylcysteine synthase, the enzyme involved in the synthesis of glutathione46. In studies carried out in cultured aortic smooth muscle cells (ASMCs) low micromolar concentrations of resveratrol induced significantly the expression of cellular antioxidants in a concentration- and/or time-dependent fashion. These enzymes include superoxide dismutase, catalase, glutathione, glutathione reductase, glutathione peroxidase, glutathione S-transferase (GST), and NAD(P)H:quinone oxidoreductase-1 (NQO1). The induction of these enzymes afforded a remarkable protection against several different ROS-inducing agents and also led to a marked reduction in intracellular accumulation of ROS47. Resveratrol increases the expression of antioxidant enzymes in muscle in an effect that seems to be dependent on activation of AMPK at least in cardiac muscle48.
Resveratrol induces vasorelaxation in muscleAnother interesting issue in aging is the decrease of blood flow due to atherosclerosis and general vascular damage. The induction of NO-dependent signalling induced by CR and exercise produces a positive effect on vasorelaxation in muscle and the organism. Actually, short-term exercise sessions already improve endothelium-dependent vasodilatation49,50 and regular exercise improves largely vasodilatation that slowly returns to sedentary levels within a week of detraining49. Exercise has also improved vasorelaxation in both normotensive and hypertensive individuals through inducing the release of NO51. Moreover, exercise has been also considered as a good therapy to decrease morbidity in patients showing cardiovascular disease by its effects on blood vessels52,53.
Polyphenols present in grape extracts have also shown to decrease low-density lipoprotein-cholesterol oxidation and platelet aggregation that ameliorates several cardiovascular diseases. However, these polyphenols also show additional cardioprotective and vasoprotective including vasorelaxation actions54. Resveratrol treatment also induces vasorelaxation in several different arteries by mechanism that seems to depend on NO availability55,56 and modifications of the activity of ion channels57,58.
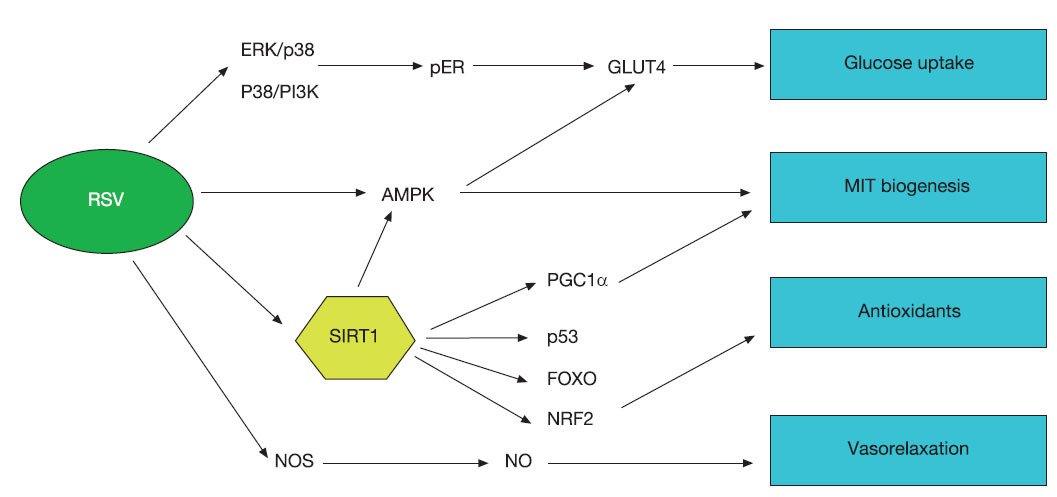
Molecular mechanisms of the activity of resveratrol in muscleThe number of pathways affected by resveratrol is currently rising severely making difficult to understand the whole picture of the effect of this compound in the organism. Figure 1 we resume the main pathways affected by resveratrol is muscle cells indicating the main function of each one although many of these pathways are involved in different physiological responses.
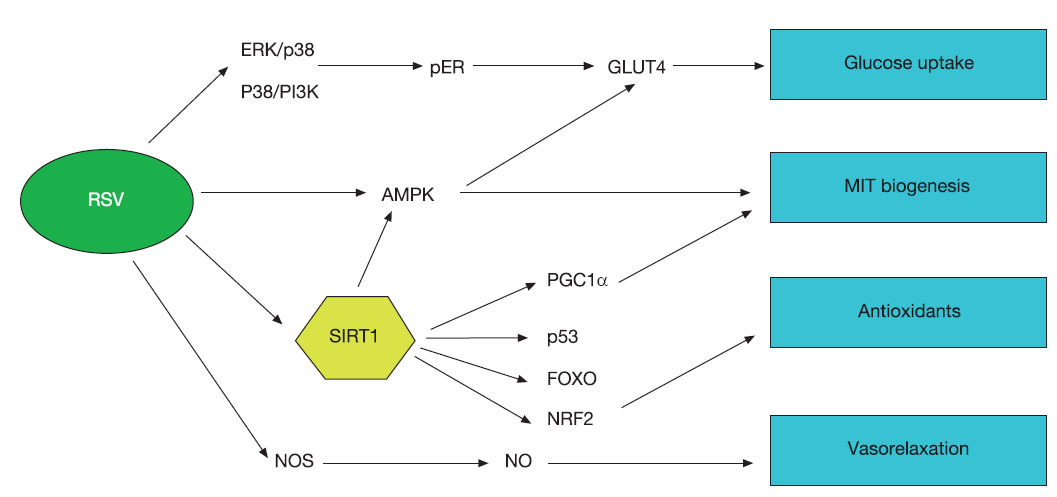
Fig. 1. Schematic signalling pathways induced by resveratrol (RSV). Currently, the number of signalling pathways induced or modulated by resveratrol is increasing. For muscle performance it has been demonstrated that resveratrol can induce glucose uptake by increasing activity or MAPK pathway and activating glucose transporters in membrane (GLUT4). Directly or indirectly AMPK is activated by resveratrol and this kinase is involved in the regulation of both, glucose uptake and mitochondrial biogenesis. The most important factor induced by resveratrol is SIRT1, that seems to be a centre component in the signalling dependent on resveratrol and regulates most of the responses to this substance by deacetylating and thus, modulating, the activity of several other components such as PGC1α, FOXO, p53, NRF2, PPARδ, etc. Several of these components are involved in mitochondrial biogenesis and then, muscle fibre switch, antioxidant enzyme induction, glucose uptake, insulin sensitivity, etc.
One of the main regulators activated by resveratrol is the NAD(+)-dependent deacetylase SIRT1. SIRT1 deacetylates other proteins and then, regulates its activity. By this mechanism, SIRT1 is a key component of the mechanism that controls metabolic processes in response to changes in nutrient availability. It has been demonstrated that activation of SIRT1 in mice is one of the key components involved in the increase or physical performance after resveratrol treatment37,38. Recently, synthetic analogues of resveratrol such as SRT1720 that shows higher activation of SIRT1 but did not affect directly AMPK have robustly enhanced endurance running performance and induce increase of type I fibres in gastrocnemius muscle in mice at the same time that protect against obesity and insulin resistance59. The main mechanism of these compounds depends on the deacetylation of PGC-1α, FOXO1, and p53 by SIRT1 and the indirect stimulation of AMPK signalling through a global metabolic adaptation mimicking low energy levels like those induced by CR or exercise59.
Furthermore, in our hands, resveratrol induces the release of NO in cultured cells (Santa-Cruz Calvo, unpublished results). NO increases in skeletal muscle after exercise60 and endurance training increases the expression of the nitric oxide synthase in human skeletal muscle61. NO-dependent intracellular signalling has been also proposed as mechanism for mitochondrial biogenesis in CR11. Furthermore, several works have shown that vasodilatation induced by resveratrol depends on increases on NO production55,56 and resveratrol-induced increases of NO release have been found in smooth muscle cells62. In consequence, it is also probable that the induction of the NO release that affects mitochondrial biosynthesis is another common mechanism for exercise, CR and resveratrol in skeletal muscle.
Another important factor in mitochondrial biogenesis that could be also induced by resveratrol in muscle and directly involved in adaptive muscle fibre transformation is peroxisome proliferator-activated receptor delta (PPARδ). It has been recently shown that overexpression of an activated form of PPARδ in skeletal muscle induces muscle fibre switch increasing the number of type I muscle fibres. Moreover, treatment of wild-type mice with agonists for PPARδ also elicits a similar type I fibre gene expression profile in muscle13. These changes results in improvement against fatigue and increase of endurance and running capacity. However, to our knowledge, no effects of resveratrol on the activity of this receptor in muscle have been described to date.
Emerging polyphenol compounds in muscle performanceThe number of polyphenols that have shown effective activity on muscle performance is increasing. Very recently, green tea polyphenols have shown activity increasing the expression of glucose transporters and insulin-signal factors in both, liver and muscle63. These polyphenols, such as (-)-epigallocatechin gallate also protect muscle in a mouse model of Duchenne muscle dystrophy and stimulate muscle toward a stronger and more resistant phenotype64. Moreover, as resveratrol, green tea catechins also protect muscle cardiovascular fibres against oxidative stress induced by ischemia/reperfusion65. Furthermore, a recent work has shown that a green tea extract enriched in catechins increases muscle performance although at the same time increases lipid oxidation66. However, a more recent work suggests that long long-term intake of catechins, together with habitual exercise, is beneficial for suppressing the aging-related decline in physical performance and energy metabolism. As in the case of resveratrol, these catechins-dependent effects are due, to improved mitochondrial function in skeletal muscle67.
Then, the study of other polyphenols on muscle performance and metabolic changes must be interesting to find a correct composition of these compounds in dietary supplements specially designed for exercise.
Conclusions and perspectivesIt is clear that a life style based on a good diet and an appropriate physical activity improves health and probably increases life-span in humans. Probably, the use of CR in the case of the competition at a high level is not adequate although it is also probable that resistance runners put themselves under CR just before competition since severe diets are coupled to higher energy expenditure due to the increase of training just before competition. Thus, a more balanced and strict diet combined with higher energy expenditure will result in CR. Polyphenols such as resveratrol or some flavonoids and their derivatives are considered as dietary antioxidants but also show ergogenic capacity and can be considered as dietary supplements in high competition.
The well-known beneficial effects of exercise and CR can be due to the capability of exercise to stress the cells and induce the cellular response in a hormetic effect. Then, a sedentary lifestyle will increase the vulnerability of the body to oxidative stress whereas a more active behaviour will enhance the protection of the organism against oxidative stress and dependent diseases including aging. We consider that the combination of a diet based on a low calorie and balanced diet and physical activity can positively affect health during aging decreasing the dependency of the individuals during this period of the life.
The practice of moderate exercise together with a balanced diet and supplementation with antioxidants can be one of the more successful therapies for increasing health in the case of cardiovascular, diabetes, metabolic diseases, cancer and also aging. In fact, very recently Galan et al35 have shown a study on aged volunteers that demonstrate that regular and moderate exercise improves cardiorespiratory function and reduces the risk of cardiovascular disease in elderly people. As in the elderly, the balance between oxidants and antioxidants is displaced to the formers, antioxidant supplementation can reduce the oxidative insult specially produced by exercise in the elderly. We propose that the use of other bioactive compounds such as resveratrol or other polyphenols able to increase antioxidant machinery in cells at the same time that can act as ergogenic compounds will increase the beneficial effects of exercise specially in those people unable to perform a proper moderate exercise.
AcknowledgementsThis work has been supported by the Spanish Ministerio de Educación y Ciencia grant DEP2005-00238-C04-04.
