





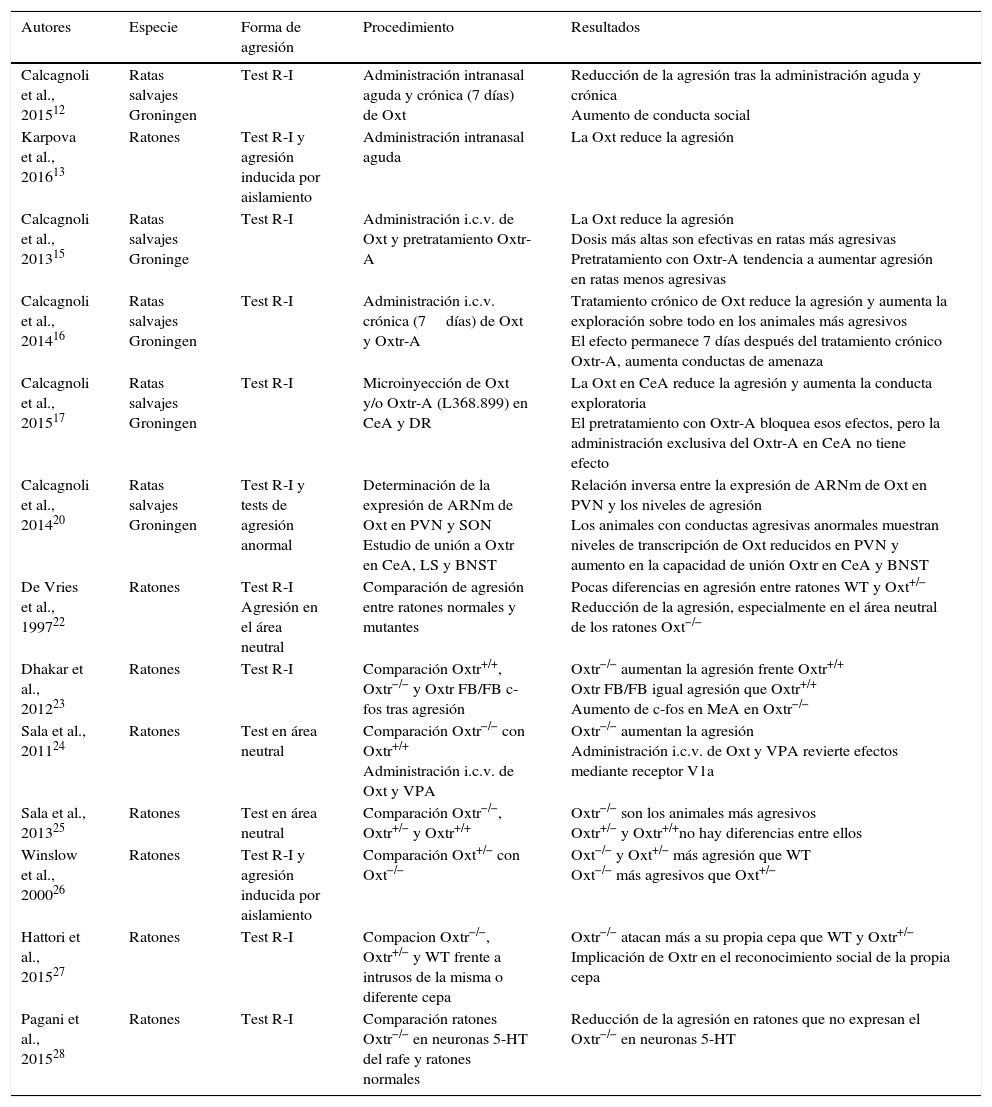
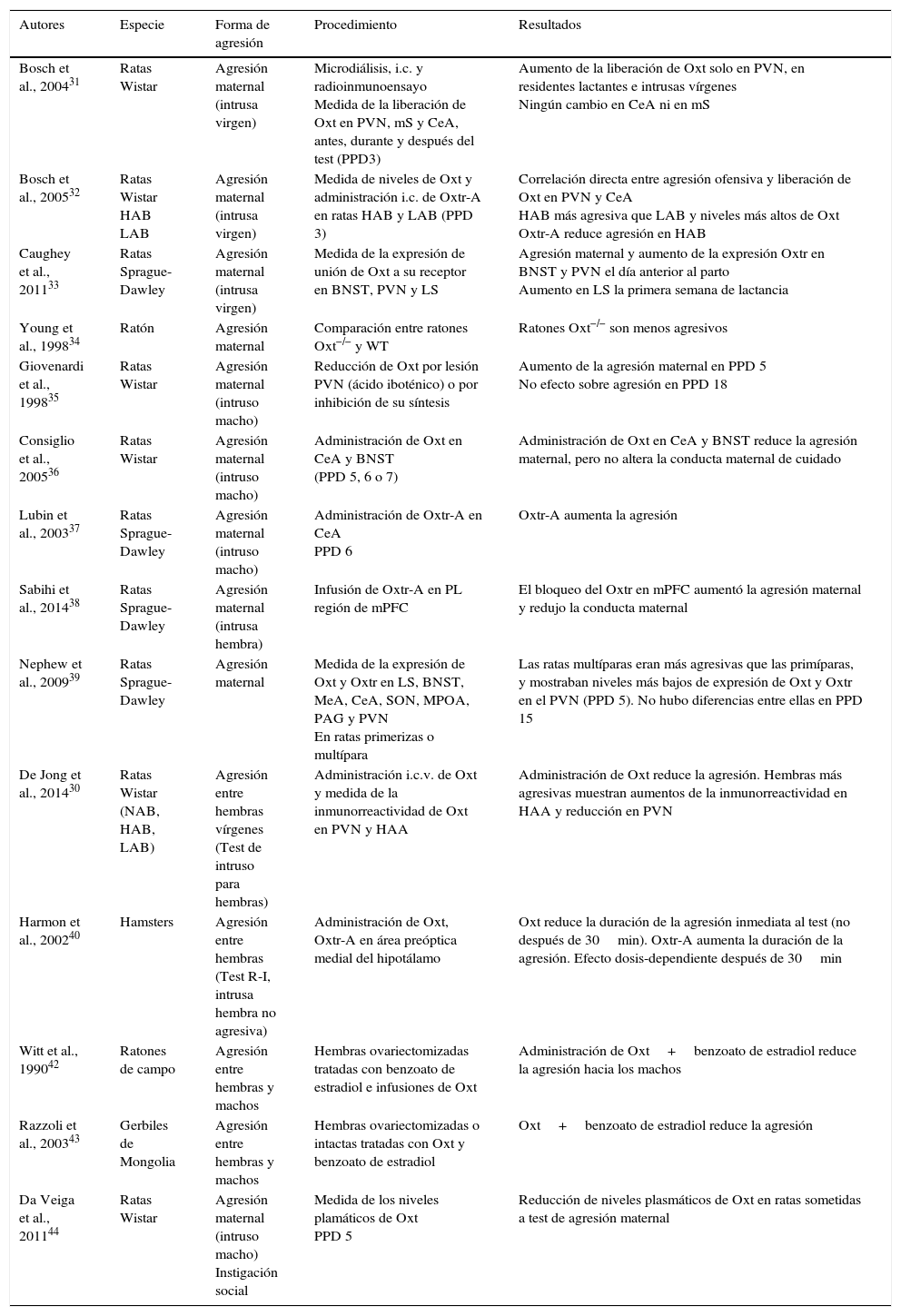

La oxitocina (Oxt) es un neuropéptido especialmente relevante por su implicación en las conductas sociales y de afiliación, incluyendo la regulación de la conducta agresiva. En este artículo presentamos una revisión actualizada que examina la relación entre la Oxt y la agresión en modelos animales. La recopilación de artículos se realizó a través de una búsqueda combinada en las bases de datos Web of Knowledge, Scopus y Pubmed (1990-septiembre 2016). Los resultados de estos estudios incluyen evidencias que proceden de la administración de Oxt intranasal, intracerebral, intracerebroventricular, o sus antagonistas, de estudios de lesión, microdiálisis, así como de trabajos con animales modificados genéticamente. En conjunto, los resultados indican que tanto en roedores machos como en hembras la Oxt modula la agresión, aunque no siempre lo hace en el mismo sentido. Se concluye que en la agresión ofensiva la Oxt ejerce generalmente un papel inhibitorio sobre la agresión, mientras que en la agresión maternal se ha observado tanto un papel facilitador como inhibidor, dependiendo de diferentes factores. Se sugiere la necesidad de analizar las variables contextuales e individuales que podrían estar modulando la acción de la Oxt sobre la agresión.
Oxytocin (Oxt) is a neuropeptide particularly relevant for its involvement in social and affiliation behaviours, including the regulation of aggressive behaviours. An updated review is presented, examining the relationships between oxytocin and aggression in animal models. The articles for review were gathered using a combined search on Web of Knowledge, Scopus, and PubMed databases (from 1990 to September 2016). The results of these studies include evidence derived from intranasal, intraventricular and intra-cerebro-ventricular Oxt administration, or from antagonist drugs, lesion studies, microdialysis, as well as works with genetically modified animals. Overall, the results indicate that Oxt modulates aggression both in male and female rodents, although it does not always act in the same direction. It is concluded that Oxt generally exerts an inhibitory role in offensive aggression, whereas in maternal aggression both a facilitating as well as an inhibitor role has been observed, depending on different factors. It is suggested that there is a need to analyse the contextual and individual variables that might be modulating the Oxt action in aggression.
Artículo
Comprando el artículo el PDF del mismo podrá ser descargado
Precio 19,34 €
Comprar ahora