


Debido a que la relación entre la masticación y las funciones cognitivas no está todavía completamente elucidada, se tuvo como objetivo determinar la influencia del tipo de masticación sobre la memoria y el aprendizaje espacial en ratones albinos machos de la cepa BALB/c.
MétodosVeinte ratones de 8 semanas de edad fueron divididos en 2 grupos iguales; el grupo masticación normal fue mantenido con alimentación en granos, la misma que recibieron los 20 ratones desde el destete; el grupo masticación deficiente se alimentó con granos pulverizados. Se evaluó a los ratones a las 16 semanas de edad durante 5 días, los primeros 4 días en la fase de adquisición y el quinto día en la fase de recuperación de memoria y aprendizaje espacial en el laberinto acuático de Morris.
ResultadosTanto en la fase de adquisición como de recuperación, no se hallaron diferencias significativas en los grupos masticación normal vs. masticación deficiente. Sin embargo, las diferencias sí fueron significativas en la fase de adquisición del grupo masticación normal, comparando los resultados del primer día vs. los otros 3 días.
ConclusionesEstos resultados indican que la masticación normal influye sobre la memoria y el aprendizaje espacial, al disminuir los tiempos de latencia en encontrar la plataforma oculta en la fase de adquisición dentro de dicho grupo experimental.
Since the relationship between chewing and cognitive functions has not been fully elucidated, this study aimed to determine the impact of chewing behaviour on spatial learning and memory in albino male BALB/c mice.
MethodsTwenty mice aged 8 weeks were divided into 2 equal groups. The regular chewing group was fed with uncrushed grains (the same diet given to all 20 mice since they were weaned) and the limited chewing group was fed with crushed grains. At 16 weeks of age, the mice were evaluated over 5 days, including a 4-day acquisition phase prior to a probe test of spatial learning and memory in the Morris water maze on the fifth day.
ResultsA comparison of the regular chewing group and the limited chewing group found no significant differences in either the acquisition phase or the probe test. However, there were significant differences in the acquisition phase for just the regular chewing group when comparing results from the first day to those from the other 3 days.
ConclusionsThe results suggest that regular chewing affects spatial learning and memory since mice in the regular chewing group decreased their times to find the hidden platform during the acquisition phase.
Las funciones cognitivas de memoria y de aprendizaje son de innegable relevancia en la vida de los seres vivos y su deterioro tiene un gran impacto social y económico en las personas, las familias, el sistema de salud y la sociedad en su conjunto1,2. Se ha encontrado que este deterioro cognitivo puede deberse a varias causas, algunas claramente entendidas, como es el caso del envejecimiento; sin embargo, también se han propuesto otras causas, no tan evidentes, que ocasionarían un declive en las funciones de memoria y aprendizaje; dentro de ellas estaría el tipo de masticación1-4.
La función masticatoria es clásicamente conocida por ser el proceso inicial de la fisiología digestiva; sin embargo, actualmente se está considerando que dicha función tiene influencias en actividades psicológicas, físicas y cognitivas1,3,5. Ampliamente ha sido demostrado que la estructura cerebral más importante implicada en las funciones cognitivas de memoria y de aprendizaje espacial es la formación hipocampal; ha sido reportado que la adecuada masticación lleva una enorme cantidad de información sensorial hacia el sistema nervioso central relacionada con el mantenimiento dichas funciones; estas vías aferentes, sobre todo, llegan a nivel hipocampal1,4-6. Encontrándose, en humanos y en animales de experimentación, que la disminución de la memoria y el aprendizaje espacial estaría asociada a una deficiencia de la función masticatoria1,4,7-9.
Por lo expuesto, se entiende que las funciones cognitivas de memoria y aprendizaje son cardinales para la adecuada supervivencia de los animales y de los seres humanos, y que las investigaciones hacen presumir que la adecuada masticación es uno de los factores que influyen en el óptimo mantenimiento de dichos procesos cognitivos. La presente investigación es el primer reporte de una serie de experimentos cuyo objetivo principal es determinar la influencia del tipo de masticación sobre la memoria y el aprendizaje espacial en ratones albinos machos de la cepa BALB/c evaluados mediante el laberinto acuático de Morris.
Materiales y métodosAnimalesSe emplearon 20 ratones machos de la cepa BALB/c adquiridos a la edad de 8 semanas de la Granja de Producción de Animales de Laboratorio del Instituto Nacional de Salud de Lima, Perú. Todos los ratones recibieron desde el destete (21 días de edad) una dieta convencional para ratón (dieta en granos). Luego de la compra de los animales, estos fueron alojados en el bioterio de la Facultad de Medicina de la Universidad Nacional Mayor de San Marcos en una habitación donde recibieran alimento y agua ad libitum, con ciclos luz/oscuridad de 12 h alternadas y con humedad y temperatura controladas (22 ± 2oC), de acuerdo con la Guide for the Care and Use of Laboratory Animals10. Después de una semana de ambientación, los 20 animales se dividieron aleatoriamente a la edad de 9 semanas en 2 grupos experimentales de 10 ratones cada uno. Para inducir la diferencia en el tipo de masticación, cada grupo experimental fue sometido durante 7 semanas a 2 tipos de dieta, los animales asignados al grupo masticación normal continuaron recibiendo la misma alimentación en granos; por otra parte, al grupo masticación deficiente se le alimentó con los granos pulverizados. Los animales fueron evaluados a la edad de 16 semanas en el laberinto acuático de Morris
AparatoEl laberinto acuático de Morris empleado fue de diseño y características similares al descrito por Dhingra et al.11 y Kitanaka et al.12, y que en resumen consistió en un tanque de plástico de forma circular de 60cm de diámetro y 30cm de profundidad, y que se dividió en 4 cuadrantes; estando colocado en una posición fija, durante todos los días que duró el experimento, dentro de una habitación (6m×4m) que tuvo características distintivas en cada una de sus paredes. El paradigma de Morris fue llenado con agua hasta la altura de 25cm, el agua se tintó con un colorante inocuo de color oscuro y se mantuvo a una temperatura de 22 ± 2oC. Una plataforma de plástico de color negro se sumergió 1cm por debajo del nivel del agua, dicha plataforma oculta tuvo un área superficial de 6×6cm y se colocó siempre en el mismo cuadrante dentro del laberinto.
ProcedimientoEl protocolo fue aprobado por el Comité de Ética de Investigación de la Facultad de Medicina de la Universidad Nacional Mayor de San Marcos (código de proyecto 0288). Los animales fueron evaluados en la fase de adquisición y en la fase de recuperación de memoria y aprendizaje espacial entre las 08:00 y las 12:00 h. La fase de adquisición se realizó durante 4 días, cada día cada ratón realizó 4 ensayos separados por 30 s de descanso; aleatoriamente se determinó cada día el orden de salida para cada ensayo dentro de los 4 cuadrantes en los que se dividió el laberinto. Un ensayo consistió en la liberación del animal con su cabeza hacia la pared de la parte central de alguno de los 4 cuadrantes posibles y se dejó libremente que nade; un ensayo concluyó cuando el animal se posó por lo menos 15 s sobre la plataforma oculta (tiempo de latencia para la fase de adquisición). El tiempo máximo de nado libre fue de 60 s; si durante dicho tiempo el ratón no encontró la plataforma, el experimentador se encargó de guiarlo hacia ella y lo mantuvo allí por 15 s, en tal caso el puntaje asignado al ratón fue de 90 s de duración del ensayo. Para esta fase, se entiende que menores tiempos de latencia en encontrar la plataforma oculta se relacionaron con una mejor memoria y aprendizaje espacial. El quinto día consistió en la fase de recuperación de memoria y aprendizaje espacial, para lo cual se realizó un único ensayo pero con características diferente del descrito; para dicho ensayo se retiró la plataforma y se liberó, desde el cuadrante opuesto adonde estuvo oculta la plataforma, a cada ratón durante 60 s, evaluándose la cantidad de tiempo que el animal nadó en el cuadrante donde estuvo ubicada la plataforma (cuadrante objetivo) y el tiempo que demoró en llegar desde su liberación hasta la zona específica donde antes estuvo ubicada dicha plataforma (tiempo de latencia para la fase de recuperación). Para esta fase, se entiende que cuanto mayor tiempo, de los 60 segundos, nadó el animal en el cuadrante objetivo y cuanto menor tiempo de latencia tuvo, se relacionaron con una mejor memoria y aprendizaje espacial. Todos los ensayos fueron filmados y luego procesados en el programa EthoVision XT 11.5 de Noldus, de donde se obtuvieron los resultados.
Análisis de los datosLos análisis estadísticos se realizaron con el programa SPSS versión 23. Los resultados se expresan utilizando la media ± desviación estándar. La normalidad fue determinada por las pruebas Shapiro-Wilk y Kolmogorov-Smirnov. En la fase de adquisición, dada la normalidad de los datos y la homocedasticidad, se utilizó la prueba t de Student, tanto para muestras independientes como para muestras relacionadas. En la fase de recuperación, al no cumplirse el supuesto de homocedasticidad, se prefirió usar la prueba no paramétrica de U de Mann-Whitney. El nivel de significación considerado fue de 0,05.
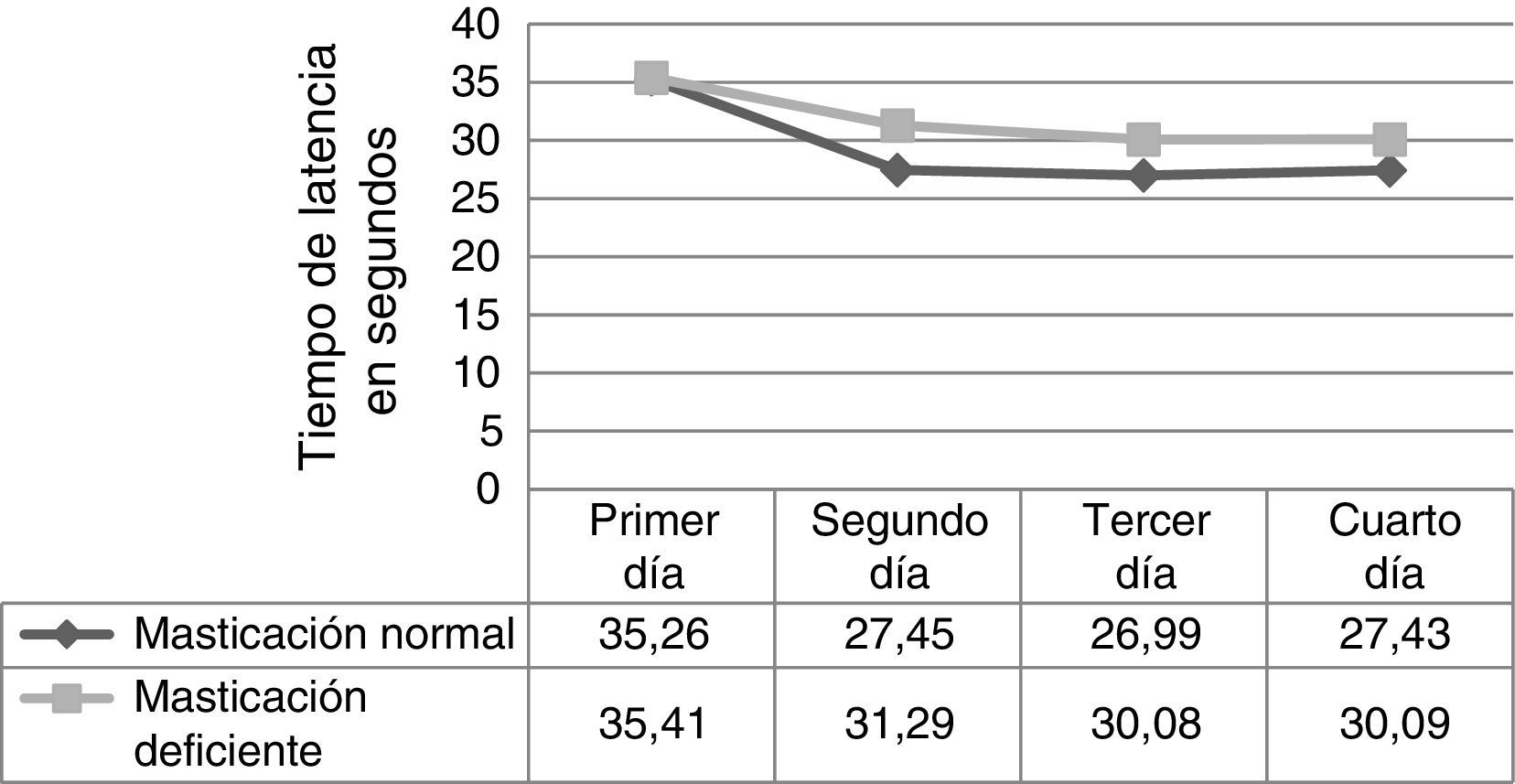
ResultadosEn relación con la fase de adquisición, se observa en la figura 1 el tiempo de latencia que empleó cada uno de los ratones para encontrar la plataforma oculta, donde se comparan las medias de los datos de los 2 grupos evaluados durante los 4 días de ensayos. Los resultados no mostraron diferencias significativas (primer día, p = 0,973; segundo día, p = 0,238; tercer día, p = 0,267; cuarto día, p = 0,317).

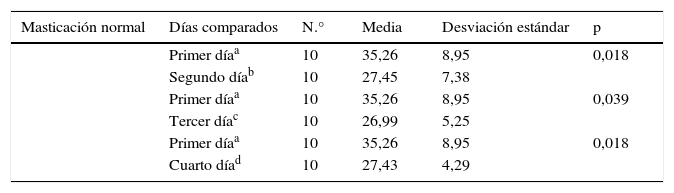
Se evaluaron los tiempos de latencia para la fase de adquisición durante los 4 días de ensayos; dentro de cada grupo, se observó que en el grupo masticación normal (tabla 1) existieron diferencias significativas al comparar el tiempo de latencia entre el primer día vs. el segundo, tercer y cuarto día; por otra parte, al comparar en el mismo grupo la latencia entre los demás días y al comparar las mismas relaciones dentro del grupo de masticación deficiente, los resultados no tuvieron significación estadística (p < 0,05).
Comparación del tiempo de latencia en encontrar la plataforma oculta en la fase adquisición en el grupo masticación normal entre el primer día vs. los demás días
| Masticación normal | Días comparados | N.° | Media | Desviación estándar | p |
|---|---|---|---|---|---|
| Primer díaa | 10 | 35,26 | 8,95 | 0,018 | |
| Segundo díab | 10 | 27,45 | 7,38 | ||
| Primer díaa | 10 | 35,26 | 8,95 | 0,039 | |
| Tercer díac | 10 | 26,99 | 5,25 | ||
| Primer díaa | 10 | 35,26 | 8,95 | 0,018 | |
| Cuarto díad | 10 | 27,43 | 4,29 |
a-d Superíndices diferentes indican resultados estadísticamente significativos.
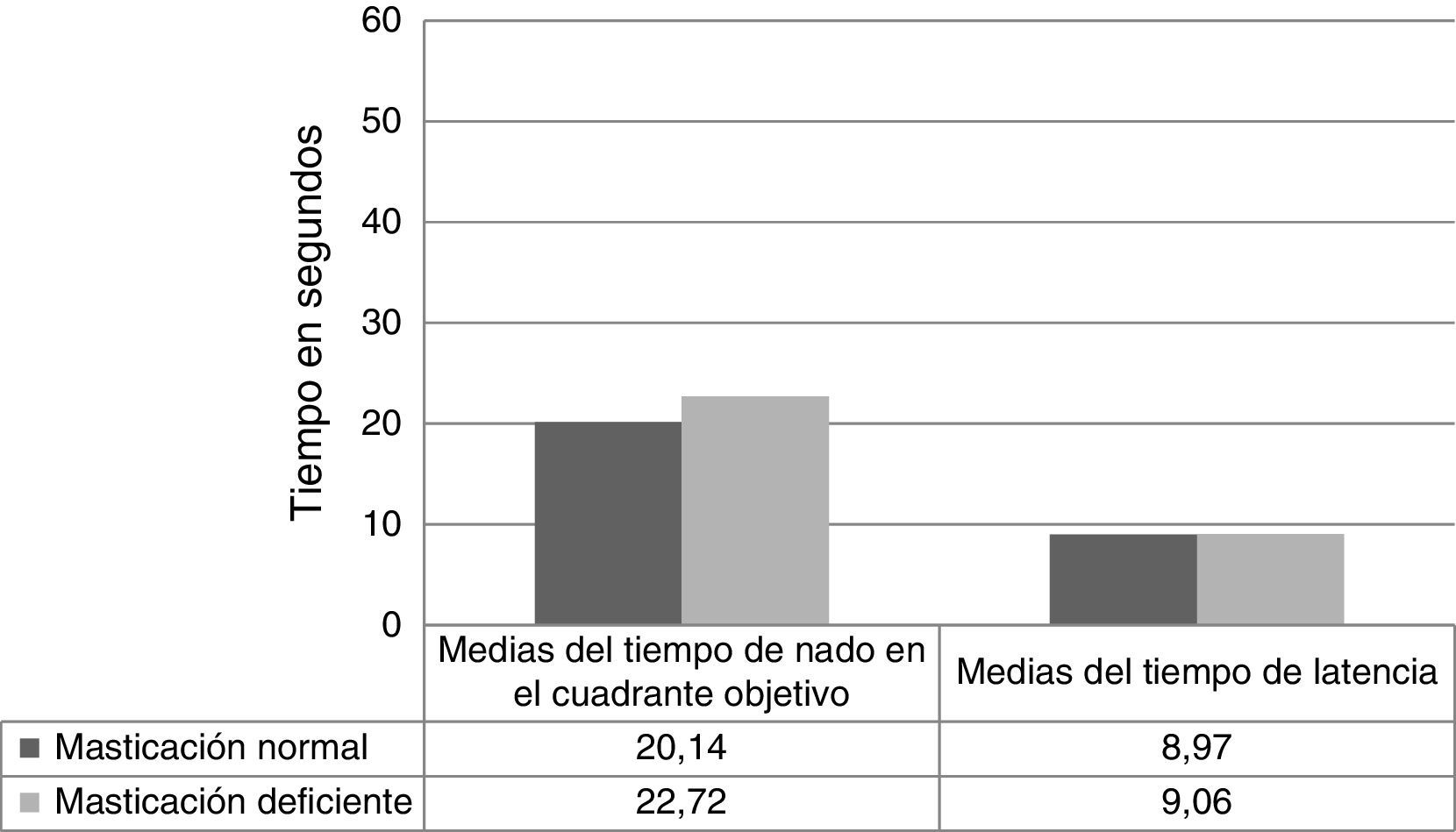
Con respecto a los datos de la fase de recuperación entre el grupo masticación normal vs. masticación deficiente, se observa en la figura 2 el tiempo que el ratón nadó en el cuadrante objetivo durante los 60 s que duró dicha prueba (20,14 ± 3,38 vs. 22,72 ± 6,09, p = 0,326) y el tiempo de latencia para la fase de recuperación (8,97 ± 3,49 vs. 9,06 ± 3,85, p = 0,910), encontrándose que las diferencias en todas las comparaciones no tuvieron significación estadística.
Discusión
El concepto de masticación normal hace referencia a un eficiente ciclo masticatorio y que depende de la actividad y la fuerza de los músculos masticatorios, de los movimientos mandibulares, de la dentición, y de la sincronía y coordinación de lengua, mejillas y labios. El tiempo que se dedique a dicho proceso determina la eficacia del mismo y dicho tiempo puede ser modulado por la consistencia de los alimentos; por otra parte, el concepto de masticación deficiente se refiere a la alteración en los factores, que fueron descritos líneas arriba, que determinan al final la pérdida de un eficiente ciclo masticatorio13, lo cual produce una inadecuada estimulación neural en el sistema estomatognático y, en última instancia, desde dicho sistema hacia el sistema nervioso central (SNC)14; así, se ha encontrado en roedores con deficiencia masticatoria una menor capacidad de memorización y de aprendizaje, produciendo dicha deficiencia cambios degenerativos y anormalidades en los mecanorreceptores periodontales, lo que indica una drástica disminución de las aferentes trigeminales desde la membrana periodontal hacía el SNC6.
Se investigó el efecto de la masticación sobre la memoria y el aprendizaje espacial en ratones. Los animales fueron divididos en 6 grupos, 3 que fueron alimentados con dieta en granos y 3 con dieta pulverizada, siendo evaluados un grupo de cada dieta a los 3, 6 y 18 meses en el laberinto acuático de Morris. Se encontró que hubo, significativamente, mejor memoria y aprendizaje espacial en los ratones con dieta en granos al compararlos, a los 6 meses de edad, con los ratones con dieta pulverizada4. Se evaluó el efecto de la masticación de goma de mascar sobre las funciones cognitivas en egresados universitarios, los cuales fueron divididos en 2 grupos iguales, uno consumió goma de mascar durante 5 minutos previos a unas pruebas cognitivas y el otro, que fue el control, no consumió goma. Los resultados encontraron que quienes masticaron goma tuvieron mejor performance que el grupo control en la batería de pruebas empleadas15. Se investigó el efecto de la masticación de goma sobre la habilidad en tareas espaciales en 100 participantes divididos en 2 grupos iguales; se emplearon pruebas seleccionados del Endless Loops Test (ELT). Los participantes del grupo experimental recibieron una goma libre de azúcar inmediatamente antes de la administración del ELT y fueron urgidos de masticar activamente durante las pruebas, el grupo control no masticó. Los resultados determinaron que no hay efectos benéficos de la masticación de goma de mascar sobre el mejoramiento en las tareas espaciales en base al ELT16. Se estudió el efecto de la alimentación con dieta blanda sobre la expresión del factor neurotrófico derivado del cerebro (BDNF) en el hipocampo. Los ratones fueron divididos en 6 grupos, 3 recibieron dieta dura (en granos) y 3 dieta blanda (pulverizada). Se sacrificó un grupo de cada tipo de dieta empleada al primer, tercer y sexto mes de vida de los animales. La alimentación blanda redujo significativamente el BDNF en el hipocampo al tercer y sexto mes, lo que correspondió a una menor cantidad de conexiones sinápticas17. El mismo grupo de investigadores evaluó el efecto de la alimentación con dieta blanda sobre la neurogénesis en el hipocampo de ratones albinos. Los animales fueron divididos en grupos que masticaron dieta blanda y dieta dura. Un grupo de cada dieta fue evaluado al tercer y sexto mes de vida. Una reducción significativa en el número de neuronas en los grupos de ratones con alimentación con dieta blanda fue reconocido en el giro dentado al tercer y sexto mes, lo que correspondió a una reducción en la neurogénesis en el hipocampo comparados con los animales alimentados con dieta dura8. Adicionalmente, hay reportes que muestran que masticar goma de mascar o el chupar un caramelo no trae ventajas en la realización de tareas cognitivas; y que el cambio de la masticación hacia la succión, en el desarrollo de actividades cognitivas, no mejora o deteriora el recuerdo18,19.
En la presente investigación, los resultados para el tiempo de latencia en la fase de adquisición fueron menores a partir del segundo día en el grupo masticación normal vs. el grupo masticación deficiente, y los resultados en la fase de recuperación fueron muy similares entre ambos grupos; sin embargo, debido a los resultados de los estadígrafos aplicados, no hay suficiente evidencia para afirmar que las diferencias observadas en la fase de adquisición se deban al tipo de masticación. Estos resultados son similares a los reportados por Frota de Almeida et al.4, quienes evaluaron en el paradigma de Morris roedores de 3 meses de edad, grupo etario que corresponde a ratones jóvenes, el mismo utilizado también en el presente trabajo, los cuales, como ocurre en el ser humano, están en una etapa de crecimiento y desarrollo hacia la plenitud de sus capacidades fisiológicas; es entendible que la masticación deficiente es sólo un factor que puede alterar la adecuada función de memoria y de aprendizaje, pero los organismos jóvenes tienen otros componentes propios de su fisiología que inciden en dichas funciones cognitivas y que compensarían la influencia de la masticación deficiente. Sin embargo, al encontrarse diferencias significativas en el grupo masticación normal al comparar los resultados del primer día vs. los demás días, se puede entender que la estimulación neural que origina la masticación normal (dieta en granos) en el ratón a largo plazo es muy importante para adquirir más rápidamente una tarea espacial al exponerlos a la misma tarea durante varios días consecutivos.
Es adecuado resaltar que en las investigaciones en roedores que han sido señaladas en párrafos anterior se maneja el esquema de dar los cambios de dieta apenas ocurre el destete y evaluar a los animales en el tiempo; el aporte de la presente investigación es que el cambio de alimentación (grupo masticación deficiente) se ha dado a la 9.ª semana de vida del animal, varias semanas después del destete, tratando de simular lo que ocurre en el ser humano, en el cual las alteraciones que originan deterioro de la masticación se van dando en el desarrollo de la vida y, generalmente, no desde el inicio de la erupción dentaria. Finalmente, sería recomendable estudiar las mismas variables en organismos de diferentes grupos etarios, pero aplicando la modificación metodológica resaltada en este último párrafo, con el propósito de continuar explorando la asociación entre las variables propuestas para posteriormente aplicar dichos conocimientos en el mejoramiento de la calidad de vida de los seres humanos, siendo el deterioro de la memoria y el aprendizaje un importante indicador en alteraciones como el deterioro cognitivo leve o la enfermedad Alzheimer, y siendo la masticación una función que, en la mayoría de los casos, es fácil de rehabilitar si es que se encuentra alterada.
FinanciaciónEl presente trabajo ha sido realizado con recursos propios del autor, lo que quisiera que se agregue es lo siguiente: El trabajo ha sido presentado como poster en el IX Congreso Internacional de Investigación del Instituto Nacional de Salud de la ciudad de Lima, Perú.
Conflicto de interesesEl autor declara que no existen conflictos de intereses en el presente trabajo.
Al Sr. Reynaldo Madrid, al Sr. César Franco Quino y al Sr. Eliberto Ruiz Ramirez.

