





El presente trabajo abarca a la alimentación de los mayas precolombinos desde tres ángulos: el perfil taxonómico de los animales consumidos, la relación entre agricultura/ecología y la obtención de los animales selváticos y la reconstrucción de algunos fragmentos de la cuisine de los mayas, basada en evidencia zooarqueológica y tafonómica. La primera parte del texto presenta listas de animales vertebrados que pudieron haber formado parte de la dieta maya, debido a que sus restos se encontraron en basureros asociados a estructuras habitacionales. A lo largo del área maya se manifiesta un patrón semejante, sobresaliendo el venado cola blanca (Odocoileus virginianus) como uno de los taxa más frecuentemente consumidos. El uso de una gama reducida pero característica de taxa que se repite con cierta constancia entre los sitios precolombinos permite relacionar la caza de animales con el uso y la transformación de la tierra en el área maya. El cultivo de la milpa parece haber propiciado la presencia de determinados taxa, importantes para el consumo. Por último, las marcas en los huesos de animales encontrados en los basureros permiten suponer que en el área maya se utilizaba, desde tiempos precolombinos, un método tradicional de cocción conocido hoy día como píib.
The present article reviews pre-Columbian Maya diet from three different angles: the taxonomic profile of the presumably consumed animals, the relationship between agriculture/ecology and animal hunting in the surroundings of the settlements, and finally, the reconstruction of some aspects of the maya cuisine, based on zooarchaeological and taphonomic evidence. The first part of the text presents lists of vertebrate animals that are presumed to have formed part of the maya diet because their remains were found in middens associated with residential structures. Throughout the maya area taxonomic studies emphasize the white tailed deer (Odocoileus virginianus) as one of the species most widely and frequently found in archaeological deposits. Also, the use of a relatively narrow range of taxa that are consistently found at most pre-Columbian sites allows us to relate the hunt of those animals to land use and transformation in the maya environment. Corn cultivation appears to have led to the increased presence of certain taxa that were important to the ancient maya diet. Lastly, the marks on the faunal bones excavated from these middens allow us to hypothesize about early evidence for a traditional cooking method, the píib, possibly used since pre-Columbian times.
La cultura maya prehispánica, ubicada en un ambiente de selvas tropicales, sabanas y manglares en la región sureste de México, Guatemala, Belice y Honduras, ha mantenido siempre un estrecho contacto con la fauna. Como resultado de numerosas investigaciones zooarqueológicas, dedicadas a estudiar la interacción entre humanos y animales en el pasado mediante los vestigios materiales excavados del registro arqueológico (véase Davis 1989), sabemos que los mayas prehispánicos interactuaron con animales domésticos y silvestres de múltiples maneras. Una importante variedad de animales cubría el lado cárnico de la dieta maya prehispánica, se intercambiaba a escala regional y/o se transformaba en instrumentos, formando bienes de prestigio, partes de vestidura, ofrendas mortuorias u otros objetos de índole cívico-religiosa.
Este artículo versa sobre la alimentación de los mayas prehispánicos con base en animales vertebrados, abarcándola desde tres ángulos y con base en una amplia revisión de la bibliografía zooarqueológica sobre el área maya. El principio básico que une a los tres ángulos es la zooarqueología. Toda inferencia que se discute en esta publicación se basa en resultados de estudios de huesos y dientes de animales vertebrados, excavados en sitios arqueológicos y analizados en laboratorios especializados para identificar su taxonomía, osteología y tafonomía.
En primer lugar se presentará un panorama amplio acerca de la variedad taxonómica de animales aprovechados en tiempos precolombinos. Discutiremos qué animales formaron probablemente parte de la dieta de los mayas prehispánicos, con base en los restos esqueléticos asociados a estructuras arquitectónicas, principalmente de función doméstica. Para esta presunción, consideramos que las acumulaciones de restos de fauna en sitios arqueológicos muy probablemente se relacionan, en principio, con desechos de la alimentación cárnica, al menos que la asociación contextual –dentro de un entierro o escondite, etc.– dicte otra interpretación. Problemáticas de interpretación que se discutirán a lo largo del texto. El segundo aspecto trata sobre los presuntos modos de obtención y caza de los animales que constituyen la parte más frecuente e importante de la dieta maya, relacionándolos con el uso y la transformación tradicionales del paisaje en el área maya. Esta sección está estrechamente relacionada con la primera, porque intenta explicar cómo y por qué dependían los mayas primordialmente de la caza de animales silvestres, mientras que hubo poco uso de animales domésticos, como en el Viejo Mundo, por ejemplo. El tercer aspecto tratado en este artículo abarca la reconstrucción de los modos de preparación de los animales que constituyeron la dieta maya precolombina, basándonos en la evidencia zooarqueológica y específicamente tafonómica de los restos esqueléticos de animales que se han encontrado en las numerosas excavaciones en el área.
La dieta maya. ¿Homogeneidad o heterogeneidad regional?Para introducir el tema de la alimentación maya prehispánica vista desde la zooarqueología, queremos someter los datos arqueofaunísticos de varios sitios precolombinos del territorio maya a una comparación con base en las publicaciones sobre el tema, para evidenciar el aprovechamiento general y, en específico, el uso alimenticio de animales vertebrados. Desde luego, debemos considerar que el material discutido en esta sección procede de los llamados “basureros”, que en el área maya consisten en acumulaciones de tiestos, fragmentos líticos, tierra y restos de animales que se encuentran asociados a estructuras arquitectónicas, o bien, constituyen incluso sus fundamentos (Chase y Chase 2000). Se presume que los basureros, cuando se hallan cerca de estructuras de uso doméstico, contienen primordialmente desechos de material de las actividades cotidianas (véase Hudson y Stanton 2006); en el caso que nos interesa en este texto, huesos y dientes de animales que reflejarían los hábitos alimenticios de los ocupantes precolombinos de las casas adyacentes. Pero un basurero puede igualmente contener desechos de la elaboración de artefactos, así como reminiscencias de algún ritual que no requiera el depósito ordenado e irreversible de los especímenes, mientras que, por otro lado, no todas las acumulaciones de restos de animales tienen que relacionarse con actividades domésticas (Emery y Brown en prensa; Stanton et al. 2008).
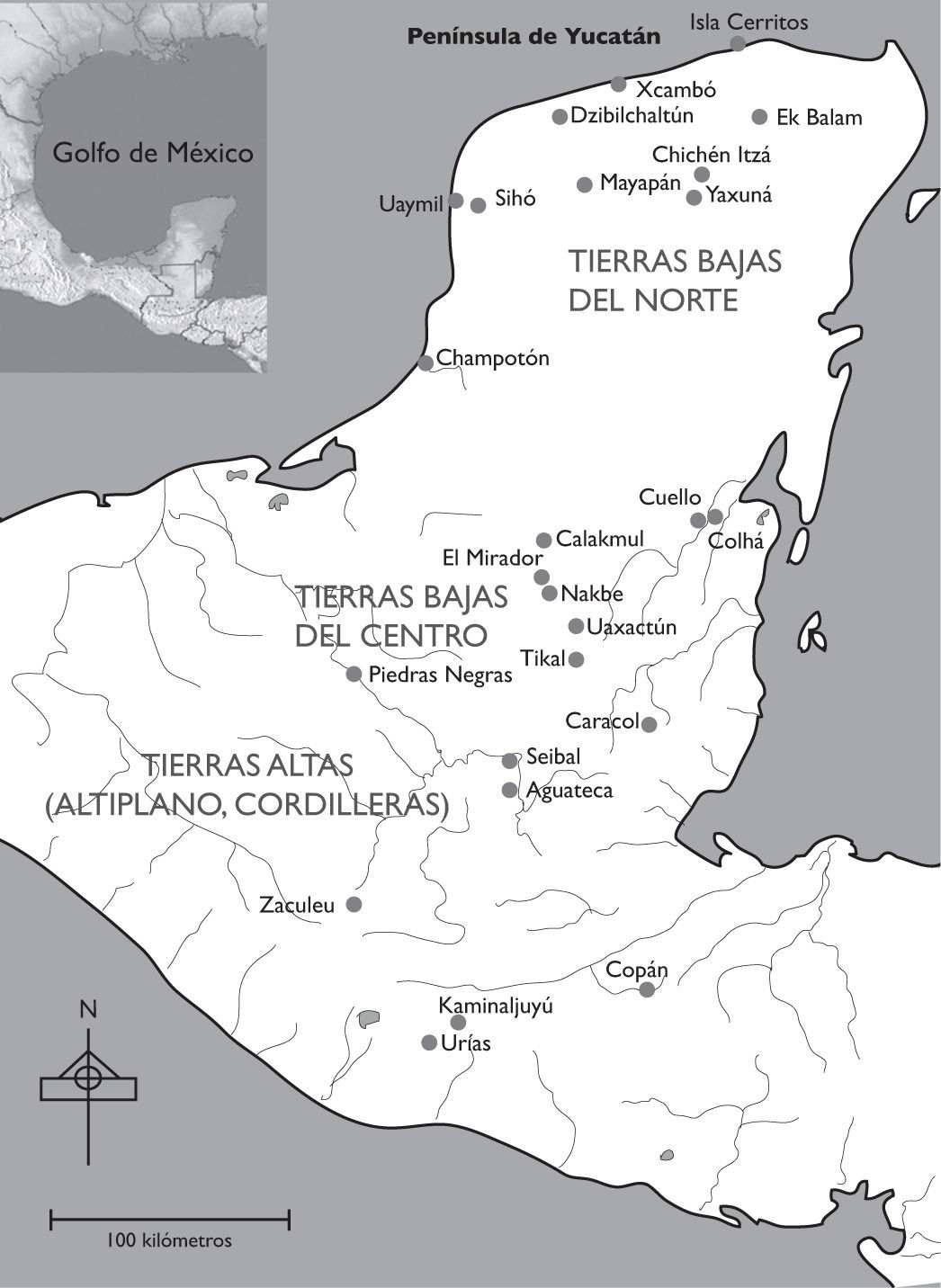
A pesar de la innegable unidad cultural que caracteriza ciertos aspectos del área maya, como la familia lingüística o los rasgos generales de la iconografía, los estilos arquitectónicos interinfluenciados o el patrón de asentamiento, se encuentran notables diferencias geofísicas, geográficas y culturales entre las tres subáreas geográfico-culturales, en las que se divide el área maya (Sharer 1994). El objetivo de la comparación es evidenciar semejanzas y contrastes que unen, por un lado, la región maya entre las Tierras Bajas del Centro (Petén), las Tierras Altas (Altiplano) y las Tierras Bajas del Norte, en relación con el uso alimenticio de la fauna y, por el otro, la subdividen (figura 1).
.")
Las reminiscencias arqueológicas de fauna discutidas en esta sección proceden, en su mayoría, de ocupaciones de los periodos Preclásico y Clásico (aprox. 800 aC-1050 dC), aunque también se dispone de algunos datos del Posclásico (aprox. 1050-1500 dC). Se presentan datos de excavaciones de estructuras habitacionales, así como de basureros de diferentes ubicaciones y asociaciones a estructuras arquitectónicas. La mayoría del material arqueofaunístico al cual haremos referencia en esta publicación estuvo asociado con actividades de la llamada élite maya precolombina (véase Chase y Chase 1992 para una definición), ya que muchas intervenciones arqueológicas se centran en las grandes y llamativas estructuras pétreas. No han sido muy frecuentes hasta la fecha excavaciones en estructuras de otros estratos sociales que no formaban parte de la élite, y por lo tanto han revelado poco material arqueofaunístico, por lo que existe un menor conocimiento respecto al uso que dichos grupos de la sociedad maya prehispánica hacían de animales (véase Emery 2003). Desde luego, una discusión sobre la alimentación de los mayas precolombinos con base en los restos de animales hallados en sitios arqueológicos depende no solamente de la asociación contextual de las reminiscencias (o sea, de factores culturales), sino también de factores tafonómicos de índole natural, que posibilitaron o no, la preservación del material esquelético. Emery (2004) discute la importancia de los factores de preservación de animales de distintas condiciones esqueléticas (animales pequeños, de huesos frágiles, en comparación con animales grandes o de huesos robustos) respecto al análisis de los modos de alimentación entre los mayas y, recientemente, se están efectuando diversos experimentos tafonómicos en la parte norte de la península de Yucatán (véase Götz en prensa).
La península de Yucatán (véase figura 1) está formada por roca caliza sin elevaciones considerables, que se extiende de las cordilleras (Tierras Altas) y del istmo de Tehuantepec en dirección noreste por unos 600 kilómetros, dividiendo así el Golfo de México y el mar del Caribe. Mientras que la superficie de la porción norte se formó entre el Mioceno-Plioceno (23-1.8 millones de años ap) y se niveló durante el Cuaternario (Bautista et al. 2003: 22), prevalecen en la parte sureña levantamientos tectónicos del Mioceno de geosuelos formados durante el Eoceno (55-34 millones de años ap), presentándose una mayor carsticidad en esta región (ibid.). Las porciones norteñas y sureñas del área maya difieren en cuanto a un gradual aumento de precipitación de noroeste al sureste (Orellana et al. 2003: 39), provocando, en combinación con el suelo y la temperatura, una flora de selvas altas perennifolias en la porción sureña, selva mediana subperennifolia y subcaducifolia en el centro y selva baja caducifolia en el noroeste (Carnevali et al. 2003: 55). Adyacente a la base de la península se encuentran las Tierras Altas, un área formada desde el Paleozoico de mucha diversidad ambiental, volcanes activos y suelos fértiles, que abarca las zonas climáticas de tierra templada y fría, a partir de los 800m y hasta más de 3000m de elevación (Sharer 1994: 26-32).
La fauna de la unidad biogeográfica del área maya es neotropical (es decir, de origen sudamericano), aunque intervienen considerables componentes neárticos (de origen norteamericano) (Arita y Vázquez-Domínguez 2003: 71; Emery 2001c). Entre las tres subáreas mayas se evidencia el llamado “efecto península” (según Simpson 1964), presentándose una reducción en la diversidad de las especies de animales desde la base de la península, y con esto desde las Tierras Bajas centrales (Petén) hasta las Tierras Bajas del norte.
Entre las especies de animales vertebrados más conocidas que se encuentran en el área maya destacan: el venado cola blanca (Odocoileus virginianus), el jaguar (Panthera onca), el manatí (Trichechus manatus), el tapir (Tapirus bairdii), la cascabel (Crotalus durissus) y la iguana negra (Ctenosaura similis), el cocodrilo de pantano y de río (Crocodylus moreletti y C. acutus) y la boa (Boa constrictor), las tortugas marinas caguama (Caretta caretta), verde (Chelonia mydas) y carey (Eretmochelys imbricata), el pavo del monte (Meleagris ocellata), el hocofaisán (Crax rubra), el quetzal (Pharomachrus mocinno) y el tucán real (Ramphastos sulfurcatus) (véase Götz y Emery 2013: índice taxonómico). Hace pocos años aún habitaban focas (Monachus tropicalis) en las costas peninsulares (Götz 2012a; Hairr 2011).
Las Tierras Bajas centrales, o Petén,1 muestran una ocupación desde el periodo Preclásico (siglos IV-I aC), con sitios importantes como Uaxactún, Nakbé o El Mirador (Clark et al. 2000). El auge en el Petén tuvo lugar durante el periodo Clásico (siglos VI-VIII dC), cuando grandes sitios, como Tikal, Caracol, Copán o Calakmul, entre otros, abarcaban extensas áreas en forma de ciudades-estado competitivas, definiendo, desde el punto de vista actual y vernáculo, nuestra representación tradicional de la cultura maya prehispánica clásica. La ocupación importante de las Tierras Bajas centrales terminó durante el fin del Clásico tardío, cuando, por diversas y discutidas causas, empezó a desintegrarse la sociedad en esta región, llevando consigo el abandono de muchos de los sitios clásicos del Petén (véase Demarest et al. [eds.] 2004, para un resumen acerca del “colapso maya”). En las últimas décadas se efectuaron varias investigaciones zooarqueológicas en el Petén, revelando datos sobre el uso de animales en el Clásico tardío. En los sitios de Copán, Piedras Negras, Aguateca, Seibal y Caracol se pudo recuperar una importante cantidad de material faunístico prehispánico. Los restos zooarqueológicos de estos sitios proceden de diferentes contextos, tanto de basureros de residencias como de escondites, entierros o cuartos abandonados, en donde los restos esqueléticos de fauna se encontraron en conjunto con artefactos y ecofactos de ocupación prehispánica.
Tanto en Copán (Pohl 1995) como en Seibal (Pohl 1978), Piedras Negras (Emery 2001a), Caracol (Giddens 2001) y Aguateca (Emery 2000), fue el venado cola blanca el animal más frecuente en los basureros asociados con las habitaciones de la élite, constituyendo entre un 50 (muestra de Aguateca y Seibal) y un 80 % (muestra de Copán) de la cantidad de especímenes óseos recuperados. Los sitios con predominancia del venado cola blanca se asemejan tanto por su posición cronológica en el Clásico tardío como por su ubicación tierra adentro, alejados del mar. El segundo lugar de frecuencia lo ocupan los perros domésticos (Canis lupus familiaris), les siguen las tortugas (Testudinae) –a excepción de Copán (Pohl 1995: 7)–, los pecaríes (Tayassuidae) y los venados temazate (Mazama spp.) (Pohl 1995, 1978; Emery 2000, 2001a). Aparentemente también se explotaron, aunque a menor escala, mamíferos medianos, como lo demuestran huesos de armadillo (Dasypus novemcinctus), tepeizcuintle (Cuniculus paca), tsereque (Dasyprocta punctata) y conejo (Sylvilagus sp.), encontrados en asociación con estructuras de la élite en Piedras Negras (Emery 2001a) y Seibal (Pohl 1978).
En los sitios estudiados se muestra una clara predominancia de restos de tortuga blanca de río (Dermatemys mawii) entre los desechos alimenticios de la élite, en comparación con otras especies de tortugas (Emery 2000, Pohl 1978: 4). De éstas, jicotea (Trachemys sp.), casquito (Kinosternon sp.), etcétera, aparecieron en contextos de arquitectura sencilla, relacionados con la alimentación de la gente común (Pohl 1978: 4). Por el otro lado, la mayoría de los perros prehispánicos de las Tierras Bajas centrales parece pertenecer a una raza común de la zona (el perro maya) (Valadez et al. 2013: 527-576); sin embargo, se encontraron evidencias de la existencia de por lo menos dos perros pelones mexicanos (xolotlizcuintli) (véase Valadez et al. 2010 para una definición) en Copán, correspondientes a la fase final del Clásico tardío (Collins 2002; Pohl 1995).
El nivel económico se refleja en la predominancia de ciertos animales que estaban, al parecer, reservados para la élite en tiempos del Clásico en los sitios peteneros (véase Emery 2003). Por otro lado, el aprovechamiento de animales en las Tierras Bajas centrales muestra diferencias socioeconómicas a través del consumo de una mayor diversidad de especies en contextos de élite –y con esto del aumento de las áreas de obtención (Pohl 1978: 5)– y también por la exclusiva presencia de determinadas partes del cuerpo en los respectivos contextos. Mientras que la élite de Seibal prefería elementos corporales de venados y jabalíes ricos en carne, como las extremidades delanteras, en los contextos de habitantes comunes se encontraron predominantemente restos del esqueleto axial y de la caja torácica (Pohl 1978: 3). En general hay una ausencia de huesos de pescado en sitios del Petén (Pohl 1990: 167), lo que indica que muy probablemente esta fuente alimenticia, a pesar de los ríos, lagunas y aguadas cercanas a los sitios, no fue aprovechada. Por el otro lado, cabe la posibilidad de que las condiciones ambientales o los factores culturales crearan efectos diagenéticos severos, que impidieron la preservación del material esquelético o no posibilitaron su hallazgo arqueológico. Es posible, por ejemplo, que los restos de pescado hayan sido quemados para producir ceniza como fertilizante para el cultivo, o bien, que los restos se arrojaran en sitios donde no fuesen de peligro para animales domésticos o niños (véase Götz en prensa; Emery y Brown en prensa).
El altiplano del área maya, que cubre regiones del centro de Guatemala y del este de Chiapas, México, muestra una ocupación significativa desde el Preclásico medio. En este tiempo empieza a desarrollarse Kaminaljuyú, el cual llega a ser el centro más importante de la región durante el Preclásico tardío y Clásico temprano debido al control ejercido sobre el yacimiento de obsidiana El Chayal (Michels 1979: 156-188; Sharer 1994: 93, 95). Analizaremos los patrones de uso faunístico en el altiplano maya a partir de material procedente de los sitios Kaminaljuyú, Urías y Zaculeu, ubicados en el altiplano de Guatemala. El material de Kaminaljuyú y Urías es del Preclásico, mientras que el de Zaculeu data del Posclásico. No se cuenta con datos de estas regiones fechados en el periodo Clásico. El material de Kaminaljuyú procede de tumbas de estructuras del centro del sitio (Kidder et al. 1946), así como de las áreas residenciales (Emery et al. 2013). Los restos de animales de Urías se hallaron en estructuras domésticas centrales igualmente, y se interpretaron en cuanto al consumo cárnico de los antiguos habitantes (Emery 2002). El material faunístico de Zaculeu procede de excavaciones en estructuras del centro del sitio (Woodbury y Trik 1953) y revela datos acerca del consumo y de ofrendas en escondites y entierros.
Los tres sitios del altiplano comparten, como ya se había visto al respecto de los materiales arqueofaunísticos de las Tierras Bajas centrales, la abundancia de huesos de venado cola blanca en los contextos, lo que revela su aprovechamiento (Woodbury y Trik 1953; Emery 1995, 2002). El segundo animal en cuanto al tamaño y la frecuencia en los contextos arqueológicos es, también en esta región, el perro. Sin embargo, la frecuencia de restos de perro en las acumulaciones arqueofaunísticas del altiplano maya es mucho más baja en Zaculeu (Woodbury y Trik 1953: 278) y Urías (Emery 2002: 4) que en las capas preclásicas de Kaminaljuyú, posiblemente porque los canes sirvieron como suplemento a la alimentación cárnica o para rituales de la élite en este último (Emery et al. 2013: 410-414). En entierros de Zaculeu y Kaminaljuyú se encontraron esqueletos enteros o parciales de perros, principalmente cráneos (Kidder et al. 1946: 155), así como en Urías, lo que hace pensar en el uso de esta parte para fines rituales (Emery 2002: 6).
Fuera de restos de venados y perros, se recuperaron reminiscencias de otros animales de talla menor: jabalíes (Tayassuidae), venados temazate, conejos (Sylvilagus floridanus), armadillos, tuzas (Orthogeomys spp.), tlacuaches (Didelphis spp), patos (Anseriformes), pavos (Meleagris spp.), loros (Amazona spp.) y tecolotes (Strigiformes), sin que estos restos se pudieran asociar claramente a una estructura o contexto en particular con base en publicaciones disponibles. Algunos de los animales mencionados podrían reflejar fauna intrusiva posterior al abandono de los sitios (véase Götz en prensa).
Las Tierras Bajas del norte cuentan con asentamientos importantes desde el periodo Preclásico (Ringle y Andrews V 1988, 1990) y vivieron el florecimiento cultural durante y posterior al auge y abandono de las grandes urbes mayas del Petén (Benavides 2001a, 2001b; Cobos 2004). El apogeo cultural de la porción norte del área maya se refleja especialmente en la extensión de las grandes ciudades mayas, como Dzibilchaltún (Andrews y Andrews 1980) y Chichén Itzá (Schmidt 1994, 2007) durante el Clásico tardío (600-850 dC) y Clásico terminal (850-1050 dC), pero igualmente en el florecimiento de las múltiples ciudades en la accidentada región llamada Pu’uc (Dunning 1990) y en las zonas aledañas. Las consideraciones acerca del aprovechamiento de la fauna vertebrada en el norte de la península de Yucatán y las Tierras Bajas del norte se basan en los sitios de Champotón, Chichén Itzá, Dzibilchaltún, Isla Cerritos, Mayapán, Sihó, Xcambó y Yaxuná, con datos provenientes principalmente de los periodos Clásico y Posclásico.
En todos los sitios de las Tierras Bajas del norte –Dzibilchaltún (Götz 2008: 154-164; Wing y Steadman 1980: 328), Chichén Itzá (Carr 1986: 3; Götz 2008), Mayapán (Masson y Peraza 2008), Sihó (Götz 2004, 2008) y Yaxuná (Götz 2008)–, el venado cola blanca aparece como uno de los animales más aprovechados para el consumo. Los restos de venados cola blanca subadultos en las muestras arqueofaunísticas de Mayapán son interpretados como evidencia de un manejo en cautiverio de estos animales por los antiguos pobladores del centro rector posclásico (Masson y Peraza 2008: 178-180). En las fuentes históricas del área maya, entre las cuales destacan los enunciados del obispo Diego de Landa sobre la vida de los mayas en el momento del contacto y las características naturales en el área de Yucatán, así como las indicaciones acerca de las condiciones naturales incluidas en las Relaciones histórico-geográficas, se encuentran repetidas menciones de la abundancia de venados en las tierras mayas. Los venados cola blanca son también hoy día presas preferidas durante la caza (Hostettler 1996; Jorgenson 1999). Aparte de los fragmentos de huesos y dientes de venado en los sitios de tierra adentro, aparecen en relativamente grandes cantidades, y sujetos a cambios cronológicos, especímenes esqueléticos de perro, pecarí de collar (Pecari tajacu), venado temazate (Mazama cf. pandora), pavo del monte y chachalaca (Ortalis vetula), así como de iguana negra; taxa menos representados incluyen una serie de mamíferos, aves y reptiles pequeños de los que no siempre se han podido encontrar evidencias contundentes de uso prehispánico (Götz 2008).
Es de suponer que, debido a los frecuentes hallazgos de restos de perro en contextos precolombinos, este animal fue muy aprovechado en tiempos preclásicos en Dzibilchaltún, sin que se pueda afirmar su uso ritual como en otras partes de Mesoamérica, ya que todos los restos esqueléticos de perro se encontraron en contextos domésticos (Wing y Steadman 1980: 328). Hacia el Clásico terminal se reduce su utilización en este sitio (Wing y Steadman 1980: 328), mientras que los habitantes de la pequeña ciudad de Sihó parecen haber acelerado su consumo como sustituto para la carne de grandes animales de caza (venados, jabalíes), cuyos restos esqueléticos se reducen en las capas tardías de los basureros del sitio (Götz 2004).
Los sitios costeros del norte de la península yucateca muestran un patrón diferente a los sitios tierra adentro. El material zooarqueológico de Champotón (Götz 2008, 2012a), Xcambó (Götz 2008: 170-179; Götz y Sierra 2011; Götz 2012b), Isla Cerritos (Carr 1987; Götz 2012b) y Uaymil (Götz s/d) confirma que el consumo de fauna se especializaba en animales de origen marino. La abundancia de restos de venado cola blanca no se observa en la costa. En cambio, una gran parte de los restos arqueofaunísticos es de tortugas marinas, entre ellas se pudieron identificar la tortuga caguama, carey y blanca (Götz 2008, 2012b). El énfasis en la tortuga marina aparece también en otros sitios costeros mayas y del área circuncaribeña (Wing 1981; Carr 1987; Frazier 2004).
A partir de las tortugas marinas, en la costa se consumían peces de gran tamaño, tiburones y posiblemente manatíes (Ibidem: 9), todos ellos obtenidos localmente en las aguas someras del Golfo de México. Hallazgos recientes de restos esqueléticos de focas (M. tropicalis) en Xcambó e Isla Cerritos pudieran relacionarse con el uso alimenticio, aunque aún no se han encontrado evidencias fehacientes de estas prácticas (véase Götz 2011c). Pocos taxa ictiológicos podrían haberse capturado en alta mar y ninguno está limitado a aguas pelágicas (Götz 2012b). El género de pez más abundante es el robalo (Centropomus), el cual puede soportar condiciones acuáticas semi-saladas (Robins et al. 1986: 128) de los estuarios cercanos a la costa. Restos de animales de tierra adentro son muy contados en contextos costeros y entre los pocos resaltan los venados cola blanca y el perro doméstico (Ibidem: 7). Los venados posiblemente se llevaban a las islas como cortes de carne (Ibidem: 24) y en forma de elementos útiles para la elaboración de artefactos, como astas y metapodios (ibid.).
Los contactos entre sitios tierra adentro y costeros del norte peninsular se manifiestan, en tiempos del Clásico tardío y terminal, mediante evidencias de un flujo de productos marinos, crustáceos y peces, desde el mar hacia, por ejemplo, Chichén Itzá, donde se han encontrado múltiples restos arqueofaunísticos de animales marinos (Ibidem: 4 y ss.; Cobos 1989). Los peces se transportaban posiblemente en forma seca y salada (Carr 1995: 3) y pueden haber representado en muchos sitios de las Tierras Bajas del norte un suplemento alimenticio, junto con cangrejos y conchas (Wing y Steadman 1980; Carr Ibidem).
Al comparar el panorama taxonómico de las subáreas, pueden discernirse algunos patrones para saber qué tan uniforme fue la dieta maya en tiempos precolombinos. En primera instancia debe hacerse hincapié en que los recursos faunísticos reflejan, en la mayoría de los casos, un aprovechamiento de las zonas inmediatas y adyacentes a los asentamientos. Esta presunción se basa en los requerimientos biogeográficos de los taxa que se han hallado en las excavaciones arqueológicas y encuentra su convalidación en un estudio isotópico que realizó Thornton (2011) sobre materiales arqueofaunísticos de varios asentamientos del Petén guatemalteco.
En básicamente todos los sitios tierra adentro del territorio maya y Mesoamérica en general, el venado cola blanca, un rumiante adaptable distribuido desde el centro-norte de EUA hasta el norte de Sudamérica (Burnie 2001; Smith 1991), aparenta haber sido una fuente muy importante de alimento (Álvarez y Ocaña 1999). En las publicaciones referidas se evidencia la abundante presencia de restos de estos cérvidos –mayormente de las partes carnosas del cuerpo– en contextos domésticos tanto en el Petén como en el altiplano y las Tierras Bajas centrales, desde el Preclásico hasta el Clásico terminal. A pesar de las diferencias ecológicas que caracterizan las tres subáreas comparadas en este trabajo, se puede suponer que los venados eran relativamente abundantes en todas las regiones, ya que el bosque secundario y las áreas de milpas constituyen el hábitat preferido de estos animales (Reid 1997). Tanto la abundancia postulada de venados –de los que inclusive pudo haber habido un manejo en cautiverio (Masson y Peraza 2008)– como el hecho de que constituye uno de los mamíferos más abundantes de la región, los hicieron apreciados por los estratos gobernantes y por los campesinos y trabajadores de los asentamientos mayas (Götz 2011a). El vínculo entre los hallazgos zooarqueológicos del periodo Clásico, la divinidad vista en los venados por grupos mayas durante la conquista española en el Petén (Carr 1996: 251) y las representaciones en códices posclásicos de las Tierras Bajas del norte (Lee 1985) muestran igualmente la amplia distribución del consumo y uso ritual de estos animales.
Tanto en el sitio preclásico de Cuello (Wing y Scudder 1991) como en el centro posclásico de Mayapán, se encontraron escondites con numerosas mandíbulas de venado, mientras que en las muestras domésticas posclásicas de Mayapán faltan proporcionalmente los elementos craneales (Masson y Peraza 2008: 176). Una desproporción semejante entre especímenes craneales y poscraneales se halló en los sitios tierra adentro de Chichén Itzá, Dzibilchaltún, Sihó y Yaxuná (Götz 2008: 319-333), lo que podría indicar que estas partes del cuerpo recibían un tratamiento específico y se depositaban en situaciones especiales (Masson y Peraza 2008: 180-181). Pohl (1990: 158) sugiere además que en el Petén las extremidades traseras de los venados cola blanca, notablemente ausentes en los contextos alimenticios de residencias de élite, fueron ofrendadas a los dioses, tal como parece mostrarse en el Códice Dresde (Lee 1985: 117), aunque también podrían haberse encontrado pocos de estos restos porque se usaban para la elaboración de artefactos (Emery 2001b). Una posibilidad de explicación para aquellos depósitos que incluyen grandes cantidades de huesos, no sólo de venado, sino de otros animales aprovechados por los mayas precolombinos, es la que exploran Brown y Emery (2008). Las autoras reportan grandes cantidades de huesos de animales en nichos pétreos de los alrededores del lago de Atitlán, Guatemala, que fueron depositados por los cazadores mayas rurales de la actualidad, en rituales de petición a los guardianes del bosque.
El segundo animal ampliamente distribuido en el área maya es el perro (véase también Emery 2004). El perro estuvo distribuido en toda Mesoamérica y desde el periodo Preclásico se le utilizó para la alimentación y rituales (Valadez 2003). Por otro lado, la frecuente presencia de restos de perro en basureros del área maya podría indicar que estos animales compartían los espacios domésticos con los humanos. Parece que hubo una mayor frecuencia de perros en sitios tierra adentro que en las Tierras Bajas del norte y centrales que en el altiplano, en las costas y sobre todo en las pequeñas islas. Esto último puede deberse a que el perro, único mamífero domesticado en Mesoamérica y morador común asociado a las viviendas humanas con aprovechamiento de desechos y materias primas, posiblemente no pudo mantenerse en “áreas de sustento difícil”, como son las islas pequeñas, con poco espacio y reducido acceso al agua (Carr 1987). En islas grandes, como Cozumel, la frecuencia de perros parece haber sido semejante a la de sitios tierra adentro (véase Hamblin 1984: 100-121).
En Copán, para el Clásico tardío, al igual que en contextos posclásicos de los sitios de Chac Mool, de la costa caribeña (Blanco et al. 1999) y Champotón (Götz 2008, 2012a), aparecieron fragmentos óseos que se atribuyen al perro pelón llamado xolotlizcuintli en el centro de México y ah bil en el área maya, el cual constituyó entre los aztecas una parte de la alimentación cárnica (Valadez 1995; véase también Valadez et al. 2010 para una amplia y detallada discusión del tema). Los huesos de estos perros son escasos en contextos arqueológicos mayas, pero la existencia de un xolotlizcuintli en las Tierras Bajas centrales durante el Clásico mostraría la amplia distribución de aquella raza, mucho anterior a la frecuente mención de dichos perros en las comunidades indígenas de Yucatán, donde fueron criados para el consumo y se apreciaban como regalo (Relaciones histórico-geográficas 1983: 78).
El jabalí aparece también ampliamente distribuido como fuente de alimento en toda el área maya. Sin embargo, se usó en mucho menor escala que los demás mamíferos grandes de la región, posiblemente porque la cacería de los defensivos jabalíes fue mucho más difícil y peligrosa que la de venados o la crianza de perros, sobre todo al efectuar una potencial caza con artefactos líticos.
Así como la presencia de los taxa señalados marcan semejanzas entre las diversas subáreas del territorio maya, también es posible percibir algunas diferencias entre las subáreas, que podrían relacionarse con diferencias en la dieta precolombina. En las áreas norteñas tenemos, ampliamente representado en el registro arqueológico, las iguanas (Ctenosaura similis). En vez de esto hubo en el Petén un consumo elevado de tortugas. Estos patrones aparentemente diferentes podrían explicarse por decisiones culturales, ya que las tortugas están igualmente presentes en las Tierras Bajas del norte, mientras que también existen grandes iguanas en el sur, aunque una especie con requerimientos y hábitos diferentes. Se encontraron restos de iguana verde (Iguana iguana) en contextos arqueológicos de Colhá, Belice (Scott 1982), correspondientes al Posclásico. Podría ser que se consumiera más iguana en el norte, porque la Ctenosaura se atrapa con mayor facilidad (hasta hoy día es frecuente ver niños en las comunidades que han atrapado a iguanas negras) que aquella del sur. Mientras que las iguanas negras son animales gregarios que habitan áreas abiertas y áridas y se encuentran frecuentemente en la inmediata cercanía de casas habitadas, las iguanas verdes viven de forma solitaria y prefieren cuerpos de agua, a los que escapan al ser amenazadas (véase Lee 2000: 191-194).
Se encontraron notables diferencias en la dieta, dependiendo de la ubicación específica de los sitios, diferencias que podrían explicarse con base en cierto determinismo ambiental. Los sitios costeros destacan por un patrón alimenticio diferente a las demás áreas, ya que allí, en vez del venado, aparece la tortuga marina como fuente ecológica principal de alimento. Las tortugas marinas llegan a considerables tamaños y pueden cazarse con relativa facilidad cuando arriban a las playas para poner huevos. Los sitios costeros aprovechaban además el manatí, y es notable que los huesos de estos animales están a menudo quemados (Carr 1987), quizás debido a un proceso de preparación que incluía la exposición a calores intensos, con el fin de extraer la grasa de la carne.
En términos generales puede apreciarse que los mayas prehispánicos aprovechaban la carne y éste es uno de los rasgos que unen el área maya en general y que se mencionaron al principio de esta publicación. La postulada dieta tradicional maya es semejante en toda el área, a pesar de hallarse diferencias entre las distintas regiones geográficas en cuanto a la alimentación, que parecen estar relacionadas con las condiciones ambientales, o bien, con los patrones culturales que prefirieron o prohibieron ciertos taxa. En tiempos prehispánicos existía un movimiento de bienes que incluía a determinados animales, como el intercambio de pescados de la costa a sitios tierra adentro y de animales de tierra adentro a la costa, pero tal vez este comercio se mantuvo a pequeña escala por falta de métodos para conservar la carne por un largo tiempo, lo que hubiera requerido el transporte de alimentos frescos a larga distancia. Una discusión detallada sobre el tema, que presenta resultados del comercio a corta distancia en la zona del Petexbatún, Guatemala, puede encontrarse en Thornton (2011).
El sistema milpero y la caza de animalesLos datos zooarqueológicos que se han expuesto en la sección anterior podrían utilizarse para interpretar no solamente la dieta, sino los particulares modos de obtención de la fauna vertebrada que seguían los mayas precolombinos. Un rasgo notable que caracteriza a muchos pueblos mesoamericanos en general y los mayas en particular es el poco uso de animales domésticos en la dieta, mientras que las plantas domésticas ocupaban un lugar preferencial en el sustento diario (véase Valadez 2003). En este sentido se observan diferencias fuertes con muchos de los pueblos prehistóricos e históricos del Viejo Mundo, de Sudamérica y de Asia, en los que la domesticación de animales desempeñaba un papel preponderante (Davis 1989; Gautier 1990). Cabe la posibilidad de interpretar los panoramas taxonómicos del área maya como formas de manejo faunístico muy específico, que puedan relacionarse con esta particularidad cultural (véase Götz 2012c).
Se ha constatado que la fauna aprovechada en los sitios costeros precolombinos de Yucatán es taxonómicamente más variada que en los sitios tierra adentro de la misma región, presentándose en los primeros una amplia gama de peces, tortugas y aves marinas, así como mamíferos acuáticos (Götz 2008). En los sitios tierra adentro prevalece una baja diversidad faunística, con énfasis en los artiodáctilos, aves de tierra y pocos reptiles. Omnipresente en la gran mayoría de los contextos, en especial en las Tierras Bajas mayas del norte, son los restos de perro doméstico, pecarí de collar, venado cola blanca, venado temazate, pavo de monte e iguana negra, especies silvestres cuyo requerimiento ecológico está relacionado con bosques secundarios y espacios agriculturales abiertos. Por otro lado, las muestras de restos arqueofaunísticos acumulados en contextos identificados como basureros de los sitios tierra adentro carecen frecuentemente de especies que prefieren bosques profundos, como los felinos (Panthera onca, Leopardus pardalis, Leopardus wiedii, etc.), pecarí de labios blancos (Tayassu pecari), venado temazate y hocofaisán (véanse, entre otros, Howell y Webb 2004; Leopold 2000; Medellín et al. 1998; Reid 1997; Terán y Rassmussen 1994).
Parece que los taxa que constituyeron la dieta maya precolombina pudieran explicarse retomando el concepto –no el perfil taxonómico– de cacería en jardines, que ha descrito inicialmente Olga Linares (1976) para Panamá y que los ecólogos Barrera-Bassols y Toledo (2005) aplicaron a la cacería campesina moderna del área maya (para una discusión detallada véase Götz 2012c). Este esquema encaja muy bien en el perfil faunístico observado por medio de los hallazgos zooarqueológicos, por lo que podría afirmarse que este sistema fue practicado también en la antigüedad en el área maya (véase también Carr 1995; Shaw 1999). Según Barrera-Bassols y Toledo (2005), los mayas yucatecos actuales obtienen la mayoría de la presa faunística silvestre mediante cacerías en los horticultivos adyacentes a los asentamientos, en las milpas y en los bosques secundarios, formados intencionalmente mediante el sistema rotativo de la milpa. Este sistema ocasiona, a través del uso de la técnica de la roza, tumba y quema, que las zonas en las que se practica el cultivo de la milpa se conviertan en un mosaico de bosques secundarios –áreas de barbecho– y espacios agriculturales (Eastmond y Faust 2006: 272; Hostettler 1996: 292; Terán y Rassmussen 1994: 278).
Las ventajas de la cacería de jardín y milpas son obvias. Los productos vege-tales y el ambiente modificado que crea dicho sistema agrícola permiten la existencia de las especies que se observan en abundancia en los perfiles taxonómicos aprovechados. La caza protege las cosechas, permitiendo así combinar el cultivo de la milpa con la obtención de carne silvestre (para una revisión detallada del tema, véase Emery y Ford 2008, así como Ford y Nigh 2009). La interdependencia entre la fauna que habita el ambiente modificado tras el cultivo de la milpa y la protección de las cosechas alcanza en la actualidad, y probablemente en tiempos prehispánicos, un estadio de manipulación indirecta de la fauna, que pudo haber sido una de las causas por las que la domesticación directa de animales no fue necesaria y nunca llegó a desarrollarse entre los mayas (Götz 2012c). Los datos zooarqueológicos apoyan en este sentido la hipótesis de un largo funcionamiento del sistema de cacería en milpa, tras su aparentemente exitosa aplicación a lo largo de muchos siglos e incluso milenios.
Los perfiles taxonómicos prehispánicos parecen indicar inclusive que en tiempos precolombinos se cazaban más enfáticamente especies de bosque secundario y espacios abiertos que en la actualidad, en la que hay registros de algunos taxa de bosques más cerrados (Jorgenson 1999). Al respecto de esto, es preciso considerar que la densidad poblacional de tiempos prehispánicos, con más de 1 000 sitios probablemente ocupados contemporáneamente durante el Clásico sólo en Yucatán (véase Garza y Kurjack 1980), redujo los bosques primarios mucho más que en la actualidad, en la que las poblaciones humanas se encuentran centradas desde la encomienda española (véase Restall 1997: 169, 172).
Desde luego, no todos los taxa que presumiblemente sirvieron para el sustento proceden de una cacería en la milpa en un sentido tan estrecho. Entre las facetas tempranas, clásicas y finales de los asentamientos, se evidencian en algunas muestras diferencias en cuanto al consumo de animales. Tanto en algunos contextos de Dzibilchaltún (Wing y Steadman 1980: 331) como en todos los basureros de Sihó (Götz 2004) se observa un cambio de un aprovechamiento regular de fauna grande en épocas tempranas (Clásico tardío) hacia el consumo de roedores, chachalacas y sobre todo iguanas hacia los momentos finales del Clásico. Estos cambios pueden haber respondido, en estos casos y siguiendo nuestra hipótesis, a expansiones del ambiente selvático, quizás por reducciones poblacionales, y con esto, de los campos milperos o de las influencias políticas que pudieron ejercer los dignatarios.
La utilización de fauna pequeña hacia el final del Clásico también pudiera estar relacionada con particularidades del contexto. En Dzibilchaltún, los restos de dichos animales de talla menor se asocian a estructuras no domésticas (Andrews y Andrews 1980: nota 331). En tiempos inmediatamente anteriores a la conquista española existían, como lo reporta Landa (2003: 130) oficios religiosos, como el de sacerdote nacom, cuya alimentación se limitaba a carne de pescado e iguanas durante los tres años de su cargo. En Chichén Itzá, por ejemplo, aparecieron huesos de iguana y pescados en conjunto, entre otros, en el tiro del osario, estructura cívico-ceremonial del centro del sitio urbano (Götz 2007: 62-68), y también en Mayapán se encontraron restos de pescado en estructuras no domésticas del centro (Carr 1995: 5). Podríamos proponer que ésta o semejantes tradiciones alimenticias empezaron a manifestarse más fuertemente hacia el Clásico terminal, y que podría usarse esta característica para reconocer más detalladamente la funcionalidad de ciertas estructuras.
Sin embargo, la posible reducción de fauna en el norte del área maya durante el Clásico terminal no se manifiesta en todos los contextos. El basurero del grupo dinástico de la serie inicial de Chichén Itzá, formado durante el final del Clásico tardío y el Clásico terminal con evidencia de uso hasta el periodo Posclásico (Schmidt 2007), contiene una gran cantidad de restos de fauna grande, predominantemente venado cola blanca (Götz 2008: 131-140). También un basurero doméstico del centro de Dzibilchaltún, del Clásico tardío y terminal (Götz 2008: 157-159) y el material faunístico de Yula, sitio pequeño y cercano a Chichén Itzá (Carr 1995), revelaron que sí hubo un fuerte aprovechamiento de animales silvestres grandes, como venados cola blanca y pavos del monte, en estos tiempos, lo que hace pensar que posiblemente no hubo una generalizada reducción de fauna grande en el Clásico tardío y terminal en el norte del área maya y la ocurrencia de los cambios necesita, en algunos casos, otra interpretación.
Cuisine maya prehispánica. Indicios zooarqueológicos de recetas milenariasEl paradigma de la zooarqueología es que los restos arqueofaunísticos encontrados en los asentamientos de la antigüedad reflejan las costumbres del uso y consumo alimenticio de animales por parte de humanos de antaño. El material arqueofaunístico descrito en las secciones anteriores procede de contextos identificados como basureros, por lo que se infiere en primera instancia que son restos de animales acumulados por seres humanos (Götz 2004, 2008; Stanchley 2004). Sin embargo, los taxa que se presentaron informan solamente acerca de la dieta antigua (siguiendo a Reitz y Wing 1999: 240), es decir, del panorama de taxa consumidos, sin que se demuestre en qué forma se guisaron o consumieron estos animales. Esta averiguación, relacionada con la cuisine prehispánica, tiene que basarse en un estudio más detallado de los restos faunísticos, destinado a revelar no solamente la taxonomía, sino la osteología y frecuencia esquelética, así como las marcas peri o post mortem en el hueso que denotan, en este caso, modos de procesamiento (véase Fischer 1995; Lyman 2001).
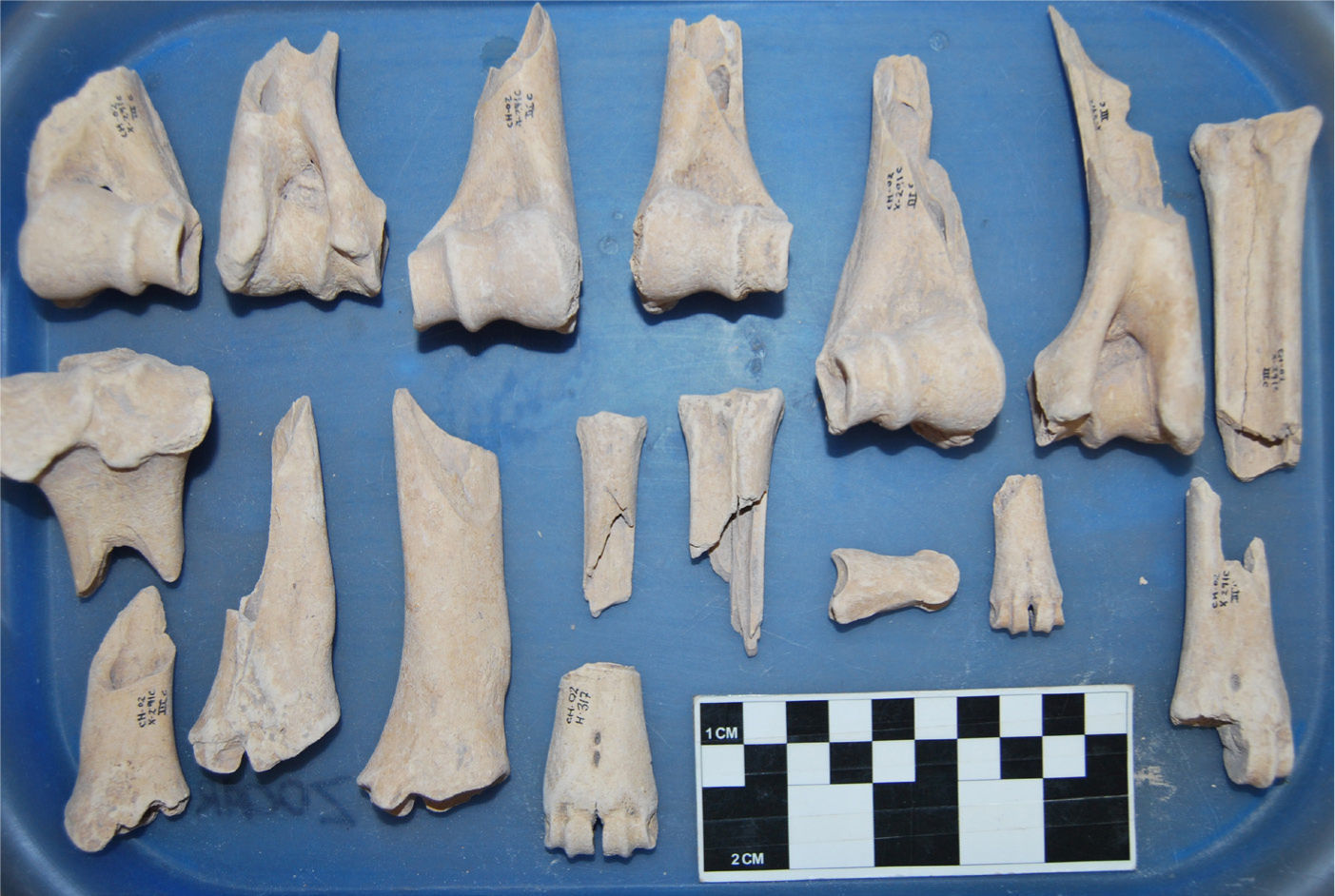
En un estudio reciente se presentaron datos de posibles modos de preparación que se seguían en tiempos precolombinos en las Tierras Bajas mayas del norte (Götz 2011b). Los datos zooarqueológicos presentados en esta publicación indican una gran cantidad de huesos arqueológicos del predominante venado cola blanca en los sitios tierra adentro con abundantes huellas de hervido y de fractura en fresco y pocas huellas de corte y de fuego directo, como se puede observar en el cuadro 1 (tomado de Götz 2011b). Las marcas de corte ocupan una posición menor en las tres muestras, mientras que la proporción de especímenes óseos con marcas de hervido y fractura en fresco es proporcionalmente alta, 28, 29, 38 y 52 %, respectivamente. La figura 2 muestra huesos arqueológicos con “fractura en fresco”, reconocible por el filo de los bordes y la forma espiral de la fractura, mismo que se relaciona con la presencia de material orgánico en el hueso córtico (véase Lyman 2001).
Frecuencia de marcas en los especímenes esqueléticos de venados (Cervidae) de los basureros de los sitios prehispánicos de Sihó (basurero de la Estructura 5D2), Chichén Itzá (gran basurero de la serie inicial) y Dzibilchaltún (basurero de la Estructura 55).
| Sitio | Cantidad total de especímenes óseos de venado | Cantidad de especímenes óseos con huellas de quemado | Cantidad de especímenes óseos con huellas de fractura en fresco | Cantidad de especímenes óseos con huellas de hervido | Cantidad de especímenes óseos con huellas de corte |
| Sihó | n=46 | n=1226% | n=24% | n=12% | n=24% |
| Chichén Itzá | n=2685 | n=1626% | n=102438% | n=78529% | n=241% |
| Dzibilchaltún | n=618 | n=8113% | n=32252% | n=17528% | n=163% |
 de venado cola blanca y venado temazate, mostrando fracturas en fresco y seco (fotografía de David Herrera).")
Se indica, además de la cantidad total de especímenes de venado por basurero, el porcentaje de huesos con huellas de fracturas en fresco, huellas de hervido y huellas de quemado, rasgos que se podrían interpretar en el sentido de procesamientos antrópicos que nos interesan en esta parte.
Los restos arqueofaunísticos prehispánicos del área maya contienen, en general, relativamente pocas marcas (Götz 2008: 83-87, 197-223), especialmente si se comparan con muestras zooarqueológicas de origen histórico de la región. Las pocas marcas de corte en relación con las fracturas en fresco y las huellas de cocción se deben, probablemente, a que los cuerpos crudos de los animales no se solían cortar en pequeñas partes durante el procesamiento (como fue presumiblemente el caso en contextos históricos), ya que parece haberse empleado, en tiempos precolombinos, un método de cocción de carne que todavía hoy día es común en las comunidades mayas rurales y que permite cocer el cuerpo antes de la partición (Götz 2011b).
Esta forma tradicional de guisar carne es el llamado píib, que es la cocción de alimentos en un horno subterráneo (Coe 1994: 158; Salazar et al. 2012). Los hornos subterráneos se construyen para la ocasión en los solares mayas, excavando un hueco en el suelo que se llena con piedras. Sobre estas piedras se enciende un fuego con abundante leña y al consumirse las llamas se colocan los guisos, envueltos en hojas y/o dentro de un recipiente, tapándolos completamente junto con las piedras, ramas y hojas de ramón (Brosimum alicastrum) y tierra (figura 3). El cocimiento de la carne se logra en los hornos subterráneos, ya que la temperatura puede alcanzar entre 200 y 300 °C. Los huesos no se ven afectados por el fuego directo, ya que en el momento de colocar el guiso se encuentra apagada cualquier flama y las partes cárnicas están envueltas con masa y hojas, formando un tamal. En experimentos realizados en torno a un proyecto de investigación en curso pudo observarse que los huesos cocidos mediante esta técnica presentan una estructura de hervido, ya que se cuecen en conjunto con los líquidos que exudan los guisos en la envoltura bajo la tierra (Götz 2013).
.")
La poca presencia de huellas de fuego parece indicar que el asado de carne no era práctica común en los sitios tierra adentro de las Tierras Bajas mayas del norte, ni tampoco el descuartizar los cuerpos antes de una preparación, mismo que dejaría marcas de desollamiento, despiece y descarnamiento. En sitios costeros parece haberse practicado con mayor frecuencia el asado, como lo atestiguan huellas de quemado en restos de manatíes (Carr 1987). También en algunos sitios tierra adentro parece que las tortugas fueron rostizadas, como lo atestiguan los fragmentos de caparazón quemados que se encontraron en varios sitios del Petén (Pohl 1990: 158). También el obispo Diego de Landa (1990: 53) describe únicamente un “asado” de carnes, al referirse a las formas de cocción seguidas entre los mayas. Esta forma de rostizar y ahumar carnes, en ocasiones previamente saladas, ya sea al colocarlas encima de una rejilla de ramas y sobre las piedras que conforman el k’óoben (el fogón tradicional mesoamericano de tres piedras), es reportada por Coe (1994: 157) como un método de cocción de carne común entre los indígenas mayas (figura 4).
 y c) Kóben tradicional maya, sobre el cual pueden cocinarse caldos, maíz o frijoles. La imagen b muestra una rejilla de ramas, utilizada para asar carnes (fotografías de Virginia Marisol Ley Lara).")
Hoy día se puede observar la cocción de tiras de carne ocasionalmente salada (de venado, jabalí, o bien, de mamíferos domésticos), ya sea colocándola a distancia de más de un metro encima del fuego para que se ahúme, o directamente sobre las piedras del fogón. Con base en los datos zooarqueológicos no podemos decir si esta técnica se aplicó en tiempos prehispánicos, ya que los huesos arqueofaunísticos no muestran marcas de corte o fileteado que evidencien el desprendimiento de la carne cruda, ni tampoco se han podido registrar frecuentes marcas de fuego y manchones negros más allá del hollín, que se hubiesen formado al colocar la carne en conjunto con los huesos adyacentes cerca del fuego. Al respecto de esto último aún falta mucha investigación por hacerse, ya que también las marcas de fuego directo que se observan en los restos arqueofaunísticos de sitios tierra adentro se podrían relacionar con daños ocurridos en momentos que se quemaba la basura o el campo al cual se habrían tirado los restos.
ConclusionesLa investigación acerca de los modos alimenticios de pueblos de antaño es un campo amplio y demandante, cuyo estudio combina varias subdisciplinas de la arqueología. En este artículo hemos tratado de reunir datos de una sola vertiente, la zooarqueología, para aportar una imagen acerca de lo que se sabe hoy día sobre la alimentación de los mayas prehispánicos con énfasis en los animales vertebrados. Se presentaron datos sobre el perfil taxonómico aprovechado, la relación presumida entre la ecología y la obtención de los taxa, y la cuisine maya con base en los conocimientos actuales. Desde luego, específicamente el estudio de las marcas relacionadas (o no) con actividades humanas es vasto e importante, ya que muchas de las interrogantes que surgen al enlistar taxa de animales presumiblemente consumidos pueden ser respondidas mediante un detallado estudio tafonómico. ¿Se consumían perros y se relacionan sus restos en los basureros con su uso culinario? ¿Realmente se rostizaban tortugas o se generaron las marcas de fuego en su caparazón debido a que estos reptiles murieron en sus intentos de escapar a los fuegos de campo? ¿Fue el píib la forma preferida de preparar carne en los sitios tierra adentro o existía una forma de preparación que deja marcas aún más sutiles? La tafonomía, el estudio de las condiciones de formación y degeneración de los contextos arqueológicos con origen orgánico, está empezando a formalizarse en el área maya y trata de responder, precisamente, estas interrogantes, para que en unos años pueda añadirse información importante a lo expuesto en estas líneas.